
Фундамент поверхностный столбчатый: Поверхностный опорно-столбчатый фундамент – заказать в Москве
Опорно столбчатый фундамент своими руками. Плюсы и минусы.
2 Ноябрь 2016 Стройэксперт Главная страница » Фундамент » Типы и виды Просмотров: 4078
Опорно столбчатый фундамент
Технологии современного строительства, промышленного и гражданского, позволяют оптимизировать характеристики возводимого строения и сократить затраты времени и денег. Для этого необходимо грамотно подойти к разработке проекта с учетом особенностей земельного участка и параметров будущего здания. Строительство опорно-столбчатого фундамента выступает одним из набирающих популярность направлений в обустройстве оснований. В чем причина успеха этого типа конструкции?
- Виды опорно-столбчатых оснований
- Опорно-столбчатый фундамент из бетонных блоков
- Опорно-столбчатый фундамент своими руками
- Возведение столбчатого основания по этапам
- Опорно-столбчатый фундамент плюсы и минусы
Виды опорно-столбчатых оснований
Главный критерий, который позволяет использовать различные варианты исполнения данного типа основания, это многообразие строительных материалов, применяемых в производстве работ.
- Условия эксплуатации.
- Сроки строительства.
- Параметры будущего строения.
- Финансовые возможности владельца.
Проконсультируйтесь с архитектором – на основе предпроектных изысканий он сможет порекомендовать вам оптимальный вариант проекта и тип материалов.
Опорно-столбчатый фундамент состоит из ряда столбов, которые выполняют из:
- кирпича,
- фундаментных блоков,
- стальных труб рассчитанного диаметра,
- труб из асбеста,
- бетона,
- натурального камня,
- деревянных бревен.
Перед планированием такого основания, необходимо провести детальные инженерно-геологические и инженерно-геодезические изыскания. Без этого комплекса работ сложно подобрать оптимальный тип конструкции и заложить фундамент необходимой прочности. Почва должна состоять из песка или супеси, а уровень залегания грунтовых вод иметь достаточную глубину.
Каждый материал накладывает на производство работ свои особенности.
Различные варианты исполнения опорно столбчатого основания
Стальные трубы обладают прочностью и устойчивостью к нагрузкам. Но имеют один главный недостаток – они подвержены коррозии и могут постепенно разрушаться под воздействие негативных факторов окружающей среды. Перед заложением опор необходимо:
- тщательно обработать поверхность труб специальными антикоррозийными средствами,
- толщина трубки должна быть не менее 4 мм,
- не допускается использование бывших в употреблении материалов – только новые трубы.
Столбы из асбеста не подвержены подобным проблемам. Также они устойчивы к воздействию воды и могут применяться на рыхлых грунтах. Но необходимо соблюдать технологию заложения и следить за качеством материала.
Бетон позволяет возводить на фундаменте сооружения средней тяжести. При этом процесс строительства простой и может быть выполнен даже не профессионалами.
Натуральный камень является «родоначальником» столбчатого основания. Он занимает большую площадь и способен выдержать большие нагрузки. Но при закладке основания важно иметь навыки кладки натурального камня – здесь есть свои особенности, требующие специальных знаний.
Дерево разрушается под воздействием влаги и насекомых. Оно имеет ряд достоинств (сопротивление деформации и движениям почвы, стоимость), но требует пристального внимания при выборе древесины и выполнении работ по заложению фундамента. Кроме того, необходимо обработать бревна специальными средствами, чтобы обеспечить сохранность его структуры.
Столбчатый фундамент имеет свои плюсы и минусы. Но, если условия позволяют заложить именно такое основание, стоит отдавать ему предпочтение. Так вы значительно сократите затраты на строительство.
к оглавлению ↑
Опорно-столбчатый фундамент из бетонных блоков
Опорно-столбчатый фундамент из бетонных блоков самый распространенный тип основания в данной категории. Его активно применяют при строительстве каркасных домов и различных хозяйственный и бытовых построек.
Его активно применяют при строительстве каркасных домов и различных хозяйственный и бытовых построек.
Опорно столбчатое основание из бетонных блоков
При строительстве небольших зданий, порядок выполнения работы следующий:
- Определите количество столбов на периметре здания. Обязательно располагать опоры в углах здания и в точках пересечения стен. Далее необходимо ставить столбы каждые 3 метра, не меньше.
- Далее выполните точную разбивку фундамента на участке с закреплением каждой опоры.
- Делаются ямы в точках расположения столбов. Глубина составляет от 15 до 30 см. Определяется от рельефа участка и характеристик грунтов.
- В каждый приямок засыпают подушки из песка, мощностью в 15 см. После поливания водой тщательно трамбуют.
- После этого делают опорные площадки для увеличения площади опоры каждого столба.
После создания нижней площадки приступают к размещению бетонных блоков. Лучше применять полнотелые блоки, имеющие большую прочность. Для скрепления блоков применяют кладочную смесь.
Для скрепления блоков применяют кладочную смесь.
Фундамент поверхностный столбчатый из бетонных блоков при правильном выполнении работ, способен прослужить длительное время и выдерживать большие нагрузки.
к оглавлению ↑
Опорно-столбчатый фундамент своими руками
После разработки и согласования проекта, можно приступать к выполнению работ на участке. Сделать опорно-столбчатый фундамент своими руками вполне возможно, хотя если у вас мало опыта в строительстве, то лучше привлечь профессиональных строителей. Важно выполнить все этапы, перечисленные ниже.
Заглубленный вариант фундамента на опорах оптимально подходит для грунтов, склонных к пучению. При этом глубину заложения устанавливают на 30 см ниже уровня промерзания грунта. Если плотность грунта на земельном участке высокая, то допускается формирование углублений под столбы в диаметр самого столба – при таком подходе, стенки ямы послужат естественной опорой. Опалубку ставят только над землей. Если грунт рыхлый, то необходимо сделать ямы в 1,5 раза шире, чтобы смонтировать опалубку для закрепления каждого столба.
Если грунт рыхлый, то необходимо сделать ямы в 1,5 раза шире, чтобы смонтировать опалубку для закрепления каждого столба.
На дно ямы закладывают обязательную подушку. Как правило, это смесь песка и гравия (щебня), которая имеет мощность до 20 см. Ее трамбуют и смачивают.
Стенки ямы, при ее использовании в качестве естественной опалубки, а также надземная часть должны быть покрыты гидроизоляцией. Это необходимо для сохранения целостности конструкции во времени. На подушку в основании следует залить немного бетона, а после затвердевания – несколько арматур небольшого диаметра – для скрепления. Каркас связывают тонкой проволокой и таким образом создают важный опорный элемент всего фундамента.
После окончательного формирования каркаса заливают бетон в каждую яму. При этом, марка применяемого бетона должна быть не ниже М200. Для приготовления раствора под опорный фундамент наиболее часто применяют состав в следующем отношении: цемент-песок-мелкий щебень в пропорциях 1-3-5.
Для выполнения работ по бетонированию основания под опорно-блочный фундамент лучше всего обратиться к специалистам – так вы обеспечите высокое качество и избежите досадных ошибок. При возведении основания будущего дома нельзя упускать даже малые детали.
Опалубка может сниматься только после окончательного затвердевания смеси. Время застывания бетона определяется множеством факторов. Среди которых, влажность и температура окружающего воздуха. Минимальный срок – 3-5 дней. Если ямы были выкопаны больше диаметра опоры, то после застывания нужно сделать обратную засыпку грунта.
к оглавлению ↑
Возведение столбчатого основания по этапам
Краткая пошаговая инструкция для строительства столбчатого фундамента.
- Выполните комплекс изысканий перед составлением проектной документации. После разработки проекта проведите точную разметку столбов по периметру будущего здания.

- Сделайте скважины (ямы) в местах установки опор. Диаметр углублений должен быть больше, чем будущие опоры – потребуется закладка гидроизоляции и опалубки.
- Трамбовка основания ямы. Это обеспечивает надежную установку опоры. Обязательно закладывать подушку для основания столба. Лучше из песка, для некоторых конструкций потребуется смесь гравия и щебня.
- И только после этого производят установку каждого столба, в зависимости от индивидуальных требований каждого материала.
В зависимости от условий конкретного участка при производстве работ накладываются индивидуальные требования.
к оглавлению ↑
Опорно-столбчатый фундамент плюсы и минусы
Опорно-столбчатый фундамент, плюсы и минусы которого широко известны и могут быть просчитаны для каждого конкретного случая, является одним из лучших типов конструкций основания на данном этапе развития.
Среди главных достоинств фундамента на опорах выделяются следующие:
- Экономия денежных средств.
 На любом строительном объекте значительную долю затрат берет на себя возведение фундамента. В случае опорного основания вы сокращаете затраты на земляные работы, объем используемых строительных материалов, время выполнения всего комплекса работ.
На любом строительном объекте значительную долю затрат берет на себя возведение фундамента. В случае опорного основания вы сокращаете затраты на земляные работы, объем используемых строительных материалов, время выполнения всего комплекса работ. - Простота конструкции. При наличии базовых знаний, вы можете сделать этот фундамент самостоятельно, даже без привлечения посторонних.
- Широкий диапазон строительных материалов, которые можно применять на объекте.
- Высокая прочность конструкции, износостойкость и долговечность.
Достоинства и недостатки фундамента на опорах
Не стоит забывать, что опорно-столбчатый фундамент обладает и некоторыми недостатками. Их обязательно нужно учитывать при выборе:
- Ограничение по весу будущего здания. Фундамент не может применяться при обустройстве крупных объектов – требуется внимательный расчет прочности конструкции.
- Отсутствие подвального помещения в здании на таком фундаменте.
- Сниженная устойчивость к опрокидыванию видов данного основания.

Очень важный совет, повторявшийся несколько раз: перед началом работ необходимо выполнить комплекс предпроектных изысканий и провести тщательный анализ и расчет с опытным архитектором.
Опорно-столбчатые конструкции находят свое применение в различных сферах строительства. С экономической точки зрения – это один из лучших вариантов основания. При этом, если соблюсти все технологии производства, то полученный результат окажется износостойким и прослужит долгое время без проблем и деформаций.
Столбчатый фундамент своими руками: пошаговая инструкция
В зависимости от того, какой грунт находится на строительном участке, приходится правильно выбирать тип фундаментной конструкции. Если грунт мягкий, подвижный, с подтоплением или с высоким уровнем пролегания грунтовых вод, тогда без столбчатого фундамента не обойтись. При кажущейся простоте проведения монтажных работ, процесс сооружения столбчатой конструкции требует точного расчета относительно несущей способности каждой опорной стойки.
Но среди всех типов основ под дом этот самый дешевый в плане расхода материала, тем более, есть возможность использовать различные стройматериалы для нее. Давайте рассмотрим, пошаговую инструкцию сооружения столбчатого фундамента своими руками.
Об одном из достоинств уже упоминалось, это малозатратное строительство. К преимуществам можно еще добавить:
- быстрое сооружение фундамента;
- нет необходимости применять строительную технику;
- простота проводимых работ, поэтому сделать столбчатый фундамент своими руками – не проблема;
- отсутствие дополнительных мероприятий, связанных с теплоизоляцией фундаментной основы;
- столбчатый фундамент легко поддается ремонту;
- можно возводить этот вид на промерзших грунтах;
- срок службы столбов при точном следовании технологии до 100 лет.
Что касается недостатков, то их не так много:
- не очень высокая несущая способность, поэтому на столбах рекомендуется сооружать легкие постройки: деревянные, каркасные, щитовые;
- устойчивость опор снижается, если они возводятся на подвижных грунтах;
- отсутствует возможность организовать подвал или подпол.

Перед тем как начинать делать столбчатый фундамент своими руками, надо ознакомиться с его классификацией, материалами и провести расчет.
Общая схема устройства
Общую схему проведения строительных работ можно обозначить несколькими главными этапами. Они, в принципе, не отличаются от возведения любого типа фундаментов.
- Проводится расчет несущей способности с учетом количества опорных столбов и веса постройки.
- Изготавливается общий чертеж столбчатого фундамента с точным указанием расстояния между стойками, сечением опор и высотою выступа над уровнем грунта.
- Проводятся подготовительные работы. Это разметка в точности по плану, земляные с выкапыванием ям — скважин, изготовлением армирующих каркасов, подсыпкой подушки в ямы, гидроизоляция и установка каркасов. Если есть необходимость, то собирается опалубка цокольной части.
- Изготовления бетонного раствора и его заливка в подготовленные скважины с последующим удалением воздуха путем штыкования смеси.
 Если в качестве материала для столбчатого фундамента используются кирпичи или блоки, то производится их укладка.
Если в качестве материала для столбчатого фундамента используются кирпичи или блоки, то производится их укладка.
Весь узел столбчатого фундамента – это объединение столбов горизонтальной лентой, которая называется ростверком. Он может быть бетонным, металлическим, деревянным или быть нижним венцом бревенчатого сруба.
Как видно, позиций по схеме не так много, поэтому и считается, что столбчатый фундамент своими руками возводить несложно.
Классификация по материалу
Как и любая основа под дом, столбчатый фундамент должен быть прочным и надежным. Поэтому для его сооружения используются строительные материалы, которые могут обеспечить необходимую прочность и надежность. К таким относятся бетон, бетонные блоки, кирпичи, камень, металл и некоторые сорта древесины, которые используются в виде бревен или бруса, установленных вертикально.
Выбор материала на самом деле ответственный момент, от которого зависит качество конечного результата. И если стоит задача – построить столбчатый фундамент своими руками, то надо правильно выбрать материал для его возведения. К примеру, считается, что монолитные конструкции лучше возводить на водо ненасыщенных грунтах, а блочные сборные на сырых.
И если стоит задача – построить столбчатый фундамент своими руками, то надо правильно выбрать материал для его возведения. К примеру, считается, что монолитные конструкции лучше возводить на водо ненасыщенных грунтах, а блочные сборные на сырых.
Монолитные железобетонные столбы
Это самая прочная и надежная конструкция. У такого фундамента под столбы высокая прочность на растяжение и сжатие. Конечно, если учесть правильно проведенное армирование.
При этом если грамотно распределить стойки по всему периметру дома, учитывая необходимое сечение опор, то на таком фундаменте можно возводить многоэтажные строения.
Добавим, что фундамент столбчатый железобетонный сооружается в тех случаях, когда необходимо залить его на достаточную глубину, и чаще ниже уровня промерзания грунта. Он прекрасно выдерживает пучение почвы, но боится соприкосновения с грунтовыми водами. Поэтому надо позаботиться о гидроизоляции опор.
Асбестоцементные столбы
Трубы из асбестоцемента для сооружения столбов для фундамента используются не часто. Хотя они прекрасно себя чувствуют в любых грунтах.
Хотя они прекрасно себя чувствуют в любых грунтах.
Несущая способность у них не высокая, но она и не требуется, потому что трубы в конструкции фундамента выполняют функции опалубки.
Для легких сооружения рекомендуется использовать трубы диаметром до 150 мм. Для тяжелых построек не меньше 250 мм.
Столбы из пластиковых труб
Здесь та же ситуация, что и с асбоцементными. То есть, пластиковые трубы выступают в роли опалубки. Они не подвергаются коррозии, не разрушаются под действием воды, поэтому для них не нужна гидроизоляция. Единственное – это надо правильно опоры залить под фундамент. А именно: установить в них армирующий каркас и залить бетонный раствор.
Из кирпичей или блоков
К кирпичным столбам для фундамента надо относиться как к самым давно известным конструкциям. Из истории европейской и российской архитектуры они являются «долгожителями».
До сих пор кирпичные столбы стоят под огромными зданиями, и стоять, как видно, будут еще не одно столетие.
Потому что отношению к выбору кирпичей в те давние времена было серьезным. Обязательно использовался жженный кирпич высочайшего качества. Поэтому, сооружая дом на столбчатом фундаменте, надо учитывать это требование.
Что касается блочных столбов, то по прочности они не уступят кирпичным, конечно, в зависимости от материала, из которого блоки изготавливаются. Предпочтение лучше отдать бетонным блокам, шлакоблок в этом плане не может нести большие нагрузки, хотя их используют под легкие постройки типа веранда, баня и так далее.
К блочным конструкциям можно отнести столбы, возведенные из камня. Неплохой вариант, достаточно прочный, но подвиги грунта такие опоры выдержать не могут. Поэтому, когда строится дом на склоне, то от каменных стоек лучше отказаться. Как замена им – фундаменты из бутобетона. Это когда в подготовленную опалубку засыпаются камни, а затем их заливают бетонным раствором.
Деревянные столбы
Такой столбчатый фундамент, как и кирпичный, используется в России давно.
Сегодня он не так востребован, но, к примеру, под бани, установленные у реки или озера, они встречаются часто.
Надо отдать должное правильному выбору древесины. Чем она прочнее, тем дольше срок эксплуатации столбов. При этом необходимо очень тщательно провести защитные мероприятия.
- Обработка всего столба антисептическими составами.
- Обработка углубленной в грунт части битумными мастиками с приклеиванием слоя или двух рубероида.
Все это сделать самому – не проблема. Главное – не только сделать защиту, но и установить под столбы деревянные опоры. Они делаются из доски толщиною не меньше 50 мм и шириною по всей площади в два раза больше, чем сечение столба. Опорные доски также надо защитить от влаги и негативного действия грунта.
В основном используют деревянные стойки под легкие сооружения, хотя еще лет сто тому назад на них строили боярские хоромы в несколько этажей.
Морозное пучение – это та сила, которая при проектировании столбчатого фундамента берется в расчет в первую очередь.
Это очень опасное давление, которое при неправильной закладке может разорвать основу дома.
Поэтому большое количество вариантов сооружения зданий рассматривается с точки зрения воздействия на фундаментную конструкцию морозного пучения.
Основное правило проектирования гласит – фундамент должен закладываться ниже глубины промерзания грунта на 30 — 50 см. И если данный показатель равен, к примеру, 1,2 м, то соответственно глубина заложения будет равна 1,5 — 1,8 м.
Но часто легкая постройка не может оказать такого давления на стойки опор, чтобы сдержать силы морозного пучения. То есть, даже при закладке на большую глубину фундамент будет подвергать деформациям. Поэтому когда строится столбчатый фундамент своими руками, надо учитывать, что существует еще две позиции: мелкозаглубленный и поверхностный.
Незаглубленный
Незаглубленные столбчатый фундамент или поверхностный – это все те же стойки, только их глубина заложения не превышает 30 — 40 см. Часто их просто сооружают на поверхности грунта, предварительно сделав подушку и гидроизоляцию. Проектировщики относят эту разновидность к тем фундаментам, глубина заложения которых не превышает треть от уровня промерзания грунта. Из нашего примера: 120 : 3 = 40 см.
Часто их просто сооружают на поверхности грунта, предварительно сделав подушку и гидроизоляцию. Проектировщики относят эту разновидность к тем фундаментам, глубина заложения которых не превышает треть от уровня промерзания грунта. Из нашего примера: 120 : 3 = 40 см.
Устройство столбчатого фундамента данного типа относится к категории самых дешевых и быстровозводимых конструкций. Как показывает практика, для них обычно используют блочную модификацию с применением блоков, камней или кирпичей. Но так как высота столбов не очень большая, а значит, их несущая способность тоже невысокая, поэтому рекомендуется увеличить площадь сечения опор. Минимальный размер – 40 х 40 см.
Мелкозаглубленный
Мелкозаглубленный фундамент из своего названия говорит о том, что он заглубляется в грунт, но не на большую глубину. Во всяком случае, не ниже уровня его промерзания. Проектировщики глубину их заложения ведут из расчета 0,5 — 0,7 от глубины промерзания земли. Опять — таки из нашего примера: 120 х 0,5 = 60 см. Основное требование к данной конструкции – не касаться грунтовых вод.
Основное требование к данной конструкции – не касаться грунтовых вод.
С ростверком
Узел столбчатого фундамента – ростверк несет на себе одну единственную задачу – распределить нагрузку от дома на все столбы равномерно. Как уже было сказано выше, ростверк может быть сооружен из бетона, металла (швеллер или двутавр), дерева (брус сечением 150 х 200 или 200 х 200 мм) или быть бревном первого венца сруба.
Пошаговая инструкция строительства своими руками
Переходим к основному вопросу, как правильно надо сделать столбчатый фундамент своими руками. Выше уже была описана схема проведения работ, где первым этапом было проведение расчетов и составление проекта. Если на столбчатом фундаменте будет сооружаться дом, то этот этап строительства лучше передать специалистам. Ведь для того чтобы точно сделать все расчеты, необходимо учесть большое количество разных показателей и нюансов. К примеру:
- тип грунта на участке;
- уровень пролегания грунтовых вод;
- уровень промерзания почвы;
- тип строения, его этажность, из каких материалов оно сооружается;
- материалы, из которых предполагается возводить сам фундамент;
- дополнительные нагрузки.

Конечно, можно воспользоваться онлайн калькуляторами, но только не для основательных домов. Для бань, веранды, сарая его использовать можно. И то никто не дает гарантии стопроцентной правильности проведенных расчетов.
Видео
Видео о том, как самостоятельно сделать столбчатый фундамент.
Разметка и земляные работы
Итак, если проект здания на руках, то в нем обязательно проектировщиками сделана привязка фундамента к местности. Это облегчает нахождение его места расположения на плоскости. Обычно привязка проводится к границам участка, с указанием расстояния от фундаментной конструкции до этих самых границ. Поэтому перед тем как разметить столбы, надо отложить от границ эти самые размеры и определить периметр постройки.
Для этого по границам здания натягиваются два ряда бечевок, которые сразу же определяют ширину фундаментных столбов. Остальное все просто:
- на глубину 20 см снимается дерн;
- размечаются места установки столбов: по углам постройки и между ними с необходимым расстоянием, которое определено проектом;
- садовым или электрическим буром в земле делаются скважины, глубина и сечение которых также определено проектом.
Надо отдать должное, что чертеж столбчатого фундамента сильно помогает в разметке. Потому что на нем четко указаны все параметры будущей конструкции.
Устройство подушки
Подушка – это слой песка толщиною 20 — 30 см. Его засыпают в скважины и утрамбовывают подручными средствами. Это может быть брус, деревянная рейка, небольшого сечения бревно. Нередко используют трубу, к концу которой приваривают металлическую пластину.
Назначение подушки с песком – отвести часть воды от опор, которая попадает в грунт. Далее на песок заливается бетонный раствор, который впоследствии станет опорой для столбчатых конструкций. Толщина бетонного слоя 10 — 30 см.
Далее на песок заливается бетонный раствор, который впоследствии станет опорой для столбчатых конструкций. Толщина бетонного слоя 10 — 30 см.
Опалубка
С опалубкой надо разобраться тщательно. Ведь многое будет зависеть от того, какой грунт располагается на строительном участке.
- Если почва глинистая, то есть, прочная, то как таковой опалубки делать нет необходимости. Глина сама, как монолит – не обваливается и не разрушается.
- Если грунт слабый песчаный, то в скважину придется установить опалубку. При этом надо учитывать, какое сечение по форме проектировщики определили: прямоугольное или круглое. Если второе, то в качестве опалубки используются трубы: пластиковые, асбестоцементные или металлические. Если первый вариант, тогда скважину придется расширять, делать прямоугольного сечения, устанавливать в них опалубку из досок или других плоских материалов. Это большой объем земляных работ, так что такую форму для заглубленных конструкций используют крайне редко.

Что касается первой позиции, то обычно внутрь шахты закладывается свернутый в трубочку рубероид. Он формирует стенки фундамента и будет в дальнейшем выполнять функции гидроизоляции. Опалубка из рубероида – обязательный атрибут.
В независимости от того, будет ли установлена опалубка в скважину или не будет, надо обязательно сформировать столб в цокольной части. А значит, здесь опалубка собирается обязательно.
Армирование
Армирование столбов проводится в обязательном порядке, ведь именно уложенная в бетон арматура дает возможность сдерживать нагрузки от сил морозного пучения.
В проекте количество арматурных стержней, их форма соединения в каркас и диаметр указываются обязательно. Поэтому арматура просто нарезается на куски нужной длиной и связывается в каркас.
Его сечение по форме может быть треугольным, квадратным или круглым. Основная задача – правильно сложить арматурные прутки относительно друг друга на необходимом расстоянии. Потом четко их связать между собой проволокой.
Потом четко их связать между собой проволокой.
После чего каркас опускается в подготовленную скважину точно посередине. После чего можно приступать к заливке бетона.
Длина прутков арматуры подрезается с учетом, что их концы будут крепиться к армокаркасу ростверка. Поэтому стержни подрезаются на 10 — 30 см длиннее. Чтобы они на этот размер торчали над цокольной частью столбов.
Заливка бетона
К заливке столбчатого фундамента надо подходить с позиции стандартной технологии. Изготавливается бетонный раствор по классической технологии:
- одна часть цемента марки М 400;
- две части мытого песка, без большого количества глиняных примесей;
- три части щебня с гранулами 5 — 40 мм.
Самое интересное, что для столбчатого фундамента нет надобности готовить большой замес. И нет необходимости проводить заливку сразу всех столбов за один день, как это обычно делается при сооружении ленточной или плитной конструкции. Достаточно рассчитать объем для одного столба, сделать замес и залить его.
Достаточно рассчитать объем для одного столба, сделать замес и залить его.
К примеру, в качестве опалубки используется труба диаметром 150 мм, и установлена она на глубину 1,2 м. Получается, что объем пустой трубы равен:
V = SxH, где S – это площадь трубы, а H – это ее длина или глубина установки. Площадь можно найти по формуле: S = πD²/4=(3,14×0,15²)/4 = 0,018 м³. Если перевести в литры, то это будет 18 л. По сути, это два ведра раствора.
Заполняются трубы бетоном, простукивая по опалубке, проштыковывают, чтобы удалить воздух. В таком состоянии столбы должны простоять 28 дней. За это время бетон наберет свою марочную прочность.
Устройство ростверка
Будем считать, что будет ростверк монолитным по проекту. Значит, под него придется сооружать опалубку из любых плоских материалов.
Она делается на весу, поэтому под нижние щиты устанавливаются подпорки из кирпича, блоков, досок, бревен и прочих материалов. Собирается опалубка сечением в прямоугольник с полным и прочным скреплением щитов между собой.
В него укладывается армирующий каркас. Обычно это две вертикальные решетки, связанные катанкой 6 мм или арматурой 6 — 8 мм. Обязательно проводится скрепление арматурного каркаса ростверка с торчащими из столбов отрезками арматуры.
Этот узел столбчатого фундамента подвергается серьезным нагрузкам. Поэтому к креплению двух арматурных конструкций надо подходить внимательно. Производится заливка бетонного раствора с утрамбовкой и штыкованием. Через 7 дней опалубка демонтируется, через 28 дней фундамент можно нагружать.
Подведение итогов
Как видите, подходить к строительству столбчатого фундамента своими руками надо с тщательным разбором всех этапов проводимых строительных процессов. Нельзя допускать просчетов в первую очередь.
Нельзя относится к разметке в большим припусками. Надо точно бурить скважины на необходимую глубину. Даже несколько сантиметров могут в крайних ситуациях сыграть свою несущую роль. Устройство фундамента – это комплексный подход к его возведению, где нет места ошибкам и просчетам.
Виды поверхностных фундаментов — Блог о строительстве
Поверхностный фундамент— самый дешевый и наименее трудоемкий, его сооружают на плотном и сухом грунте для легких построек (небольшого дома, сарая, летней кухни, туалета и др.).
В качестве такого фундамента служат опорные плиты (рис. 8). Песчаная подушка под ними не дает образоваться льду и обеспечивает равновесие.
Рис. 8.
Поверхностный фундамент:1 – опора поверхностного фундамента; 2 – опорная плита; 3 – нижний венец постройки; 4 – забирка; 5 – песчаная подушка; 6 – зона промерзающего грунта; 7 – зона непромерзающего грунта Чтобы плита не просела, ее размер нужно рассчитать, исходя из массы постройки, допуская удельную нагрузку на плиту (для суглинистой почвы) не более 1,5—2 кг/см2.Например, при массе дома в 50000 кг, распределенном на 16 опор, размер плиты будет 50000 / 16 х 2,0 = 1600 см*, т. е. 40 х 40 смПлиты можно изготовить на месте из жирного бетона с добавлением гравия, заложив железную (проволочную) арматуру.Толщину плиты берут не менее 10 см.
е. 40 х 40 смПлиты можно изготовить на месте из жирного бетона с добавлением гравия, заложив железную (проволочную) арматуру.Толщину плиты берут не менее 10 см.
Можно применить и готовые плиты. Перед укладкой песок смачивают и трамбуют.Массу здания можно подсчитать, складывая массы затраченной древесины, кровли, кирпича для опор, снегового покрова на крыше и т. д.
Для расчетов можно принять, кг/м3: массу древесины 700—720 (ель, сосна), кирпичной кладки — 1600, снегового покрова на крыше — 500, массу кровли — исходя из массы листового железа, рубероида или шифера.Кирпичная кладка между опорами (забирка) для летних построек необязательна, так как летом под полом циркулирует теплый воздух, а зимой в летних постройках не живут.Лучший вариант — декоративная стенка между опорами. Однако, если кирпичную или бетонную стенку между опорами все же выполняют, ее не связывают с опорами фундамента. Углубляют забирку не более, чем на 15 см, укладывая ее в песчаный ров.Ещё один менее трудоёмкий и затратный ленточный поверхностный фундамент упрощенного типасооружают для легких построек — маленького садового домика, гаража, сарая и т.
п.Выкопанную траншею, заполняют слоями крупного песка, смешанного со щебнем или галькой. Каждый слой проливают водой и тщательно трамбуют. Когда траншею засыплют таким образом доверху, песчаную смесь смачивают водой и обильно поливают жидким цементным раствором.По ленточному фундаменту располагают цоколь домика.Его выполняют из того же материала, что и фундамент, или из другого, но не меньшей прочности.
По бетонному фундаменту можно сделать такой же или кирпичный цоколь, по кирпичному — кирпичный и т. д.Цоколь выкладывают поверх слоя гидроизоляции на цементном или цементно-известковом растворе. Поверх цоколя снова делают гидроизоляцию из 2—3-х слоев рубероида, приклеивая его битумом или битумной мастикой.Далее – Гидроизоляция фундаментаТакже рекомендуем: Брусчатые стеныСтены из природного камняДанный тип фундамента называют также плитным, плавающим или незаглубленным.
Поверхностный фундамент, как и все вышеописанные типы, широко применяется в индивидуальном жилом и дачном строительстве, обладая преимуществами по стоимости и простоте устройства.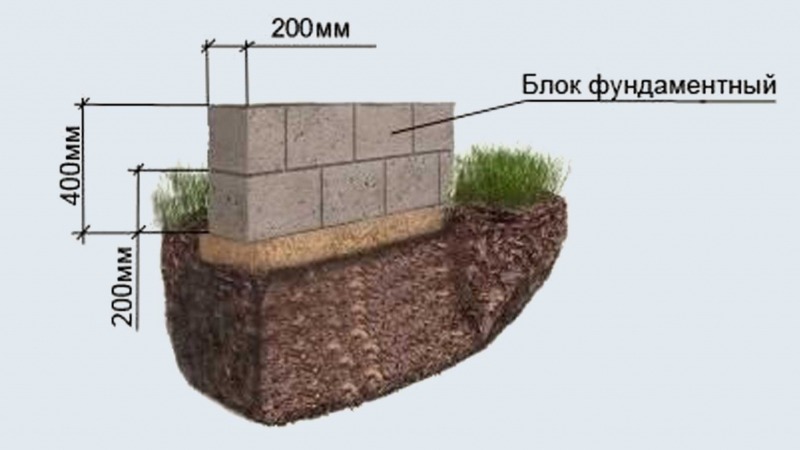 Несмотря на то что все названия этого типа фундамента являются синонимами, по условиям возведения они отличны друг от друга.Плитный фундамент можетвыполняться и заглубленным (с предварительной разработкой котлована), если необходимо устройство подвала или того требуют геологические условия. В индивидуальном строительстве заглубленный плитный фундамент применяется редко из-за своей дороговизны (поскольку на стоимость повлияют объемы земляных работ и большее по сравнению с ленточным фундаментом количество бетона и арматуры).Поверхностный плитный фундамент, устроенный на подвижном пучинистом грунтовом основании, называется плавающим, поскольку перемещается вместе с грунтом без причинения вреда конструкциям дома. Плавающие поверхностные фундаменты могут выполняться как в форме сплошной плиты, так и в виде решетчатой структуры (монолитной или из сборных перекрестных железобетонных балок).
Несмотря на то что все названия этого типа фундамента являются синонимами, по условиям возведения они отличны друг от друга.Плитный фундамент можетвыполняться и заглубленным (с предварительной разработкой котлована), если необходимо устройство подвала или того требуют геологические условия. В индивидуальном строительстве заглубленный плитный фундамент применяется редко из-за своей дороговизны (поскольку на стоимость повлияют объемы земляных работ и большее по сравнению с ленточным фундаментом количество бетона и арматуры).Поверхностный плитный фундамент, устроенный на подвижном пучинистом грунтовом основании, называется плавающим, поскольку перемещается вместе с грунтом без причинения вреда конструкциям дома. Плавающие поверхностные фундаменты могут выполняться как в форме сплошной плиты, так и в виде решетчатой структуры (монолитной или из сборных перекрестных железобетонных балок).
В последнем случае необходима жесткая заделка узлов (мест стыков балок). Наибольшая жесткость плавающего фундамента достигается устройством плиты из монолитных перекрестных лент. Плавающие фундаменты можно возводить на грунтах, подверженных сильной просадке, на насыпных грунтах (песчаная подушка, слежавшаяся свалка), на сильно пучинистых грунтах.
Плавающие фундаменты можно возводить на грунтах, подверженных сильной просадке, на насыпных грунтах (песчаная подушка, слежавшаяся свалка), на сильно пучинистых грунтах.
Для плавающих фундаментов необходимо подбирать высокопрочный бетон и арматуру диаметром не менее 10 мм.Незаглубленный фундамент сооружается не только в виде монолитной плиты, способен быть плавающим или стационарным (грунты не смещаются, поэтому и фундамент не двигается).Поверхностный фундамент по смысловому значению почти полностью совпадает с незаглубленным, однако последний может быть также ленточным или столбчатым (хотя на практике такое встречается редко).Поверхностный фундамент выполняется в виде сплошной плиты (чаще из монолитного железобетона или железобетонных блоков с обязательным армированием по всей площади и высоте). Причем земляные работы ограничиваются выравниванием площадки и разработкой неглубокого котлована.Перед тем как выбрать данный тип фундамента, обратите внимание на следующие моменты. 1. Устраивать незаглубленный фундамент нужно только с учетом прочности основания (оно может быть пучинистым, но ни в коем случае не подверженным оползневым явлениям) и всех возможных деформаций. Причем надземные конструкции дома учитываются не только при расчете нагрузки на основание, но и при определении жесткости постройки: чем она выше, тем незначительнее будет деформироваться грунтовое основание и меньше риска при устройстве поверхностного фундамента.2. Стоимость сооружения поверхностного плитного фундамента повышается за счет большого расхода материалов (ведь фундамент устраивается под всей площадью здания), поэтому целесообразно применять этот тип фундамента при возведении простых в плане зданий.
1. Устраивать незаглубленный фундамент нужно только с учетом прочности основания (оно может быть пучинистым, но ни в коем случае не подверженным оползневым явлениям) и всех возможных деформаций. Причем надземные конструкции дома учитываются не только при расчете нагрузки на основание, но и при определении жесткости постройки: чем она выше, тем незначительнее будет деформироваться грунтовое основание и меньше риска при устройстве поверхностного фундамента.2. Стоимость сооружения поверхностного плитного фундамента повышается за счет большого расхода материалов (ведь фундамент устраивается под всей площадью здания), поэтому целесообразно применять этот тип фундамента при возведении простых в плане зданий.
Однако при составлении сметы надо учесть и небольшую стоимость земляных работ, и общий объем фундамента (а не только его площадь).3. Из-за большого расхода материалов плитный фундамент чаще применяется в небольших зданиях, где сама плита фундамента способна служить полом (гараж, баня).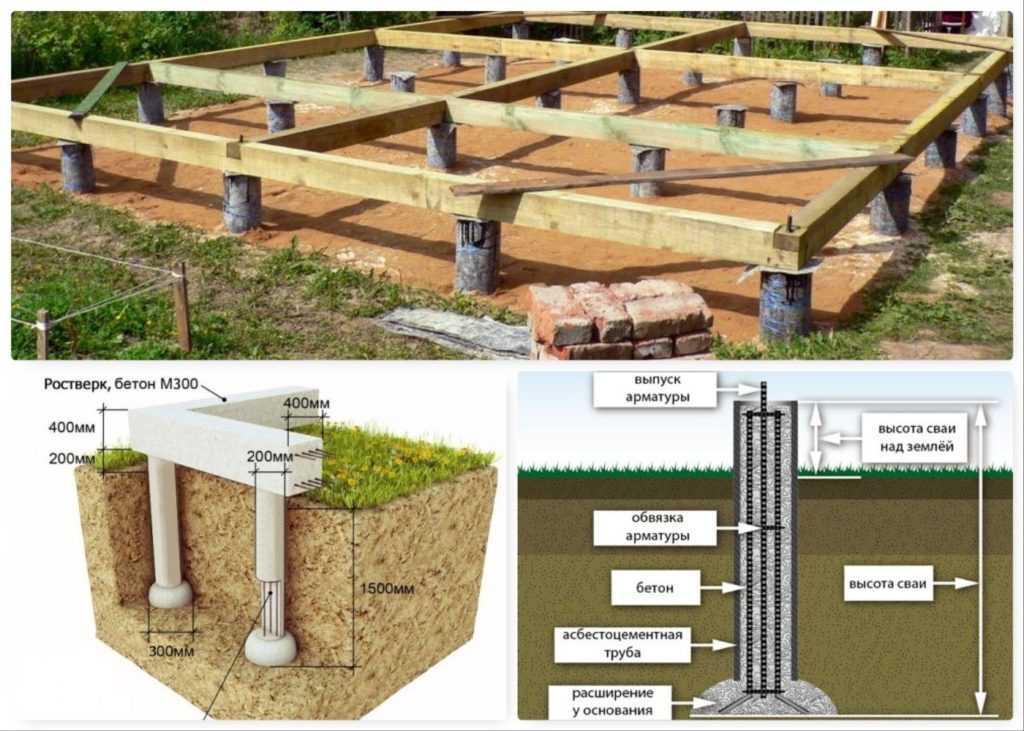 Но если сделать поверхностный фундамент не в виде сплошной плиты, а в виде ребристой монолитной плиты или армированных перекрестных лент, то такой тип фундамента будет хорошим вариантом и для жилых домов высокого класса.Плитный фундамент в здании с подвалом может обойтись дороже, чем ленточный, поэтому часто застройщики возводят ленточный фундамент, а пол цокольного или подвального этажа устраивают отдельно.
Но если сделать поверхностный фундамент не в виде сплошной плиты, а в виде ребристой монолитной плиты или армированных перекрестных лент, то такой тип фундамента будет хорошим вариантом и для жилых домов высокого класса.Плитный фундамент в здании с подвалом может обойтись дороже, чем ленточный, поэтому часто застройщики возводят ленточный фундамент, а пол цокольного или подвального этажа устраивают отдельно.
Однако подобная конструкция в отличие от плитного фундамента не обеспечивает надежную гидроизоляцию и больше подвержена просадкам.4. Данный тип фундамента обладает существенным достоинством: если конструкция его жесткая (монолитная плита), то при выполнении такого фундамента под всей площадью здания на его конструкции не будут существенно влиять перемещения грунта (как сезонные, так и любые другие). Фундамент станет двигаться вместе с грунтовым основанием (отсюда и название «плавающий»), поэтому конструкции не будут деформироваться и трескаться.Но именно поэтому грунтовое основание не должно быть очень подвижным (иначе дом может в буквальном смысле слова «уползти» от вас). По отношению же к пучинистости (воздействию замораживания, оттаивания и просадки грунта) такой фундамент вполне устойчив.5. Сооружение плитного поверхностного фундамента целесообразно на грунтовом основании с высокой степенью пучинистости и просадки (вертикальная подвижность грунта сильная, а горизонтальная — умеренная).6.
По отношению же к пучинистости (воздействию замораживания, оттаивания и просадки грунта) такой фундамент вполне устойчив.5. Сооружение плитного поверхностного фундамента целесообразно на грунтовом основании с высокой степенью пучинистости и просадки (вертикальная подвижность грунта сильная, а горизонтальная — умеренная).6.
Если требуется высокий цоколь, лучше остановить выбор на ленточном фундаменте.Когда же достаточно невысокого цоколя, верх плитного фундамента способен послужить цокольным перекрытием.От степени пучинистости грунта зависят толщина плиты основания и способ выполнения плитного фундамента:— сильное пучение предполагает устройство фундамента из монолитного железобетона или готовых блоков, жестко скрепленных между собой;— пучение средней силы позволяет возводить фундамент из монолитного железобетона или блоков, уложенных в перевязке на растворе;— если пучение слабое, но все же присутствует, подойдут цементогрунт, бутобетон, бут, керамзитобетон.Минимальная толщина плитного фундамента — 10 см. Чаще применяется плита толщиной 20—25 см, поскольку дальнейшее увеличение толщины приведет к росту нагрузок на слабый грунт, а применение минимальной толщины (10 см) не удобно для армирования и бетонирования (как следствие — понижение качества работ).Для закладки плитного фундамента необходимы те же материалы и инструменты, что и для монолитного ленточного фундамента.Требования к арматуре более высокие, чем в других типах фундамента. Она должна быть обязательно ребристого сечения, диаметром 12—16 мм.Устройство поверхностного фундамента происходит в следующей последовательности.Земляные работы: выработка котлована, выравнивание площадки, трамбовка поверхности, устройство подушки из слоя песка и слоя гравия.Укладка на подушку гидроизоляционных материалов.Заливка поверху гидроизоляции тонкого слоя бетона из подвижной смеси.Укладка арматуры.
Чаще применяется плита толщиной 20—25 см, поскольку дальнейшее увеличение толщины приведет к росту нагрузок на слабый грунт, а применение минимальной толщины (10 см) не удобно для армирования и бетонирования (как следствие — понижение качества работ).Для закладки плитного фундамента необходимы те же материалы и инструменты, что и для монолитного ленточного фундамента.Требования к арматуре более высокие, чем в других типах фундамента. Она должна быть обязательно ребристого сечения, диаметром 12—16 мм.Устройство поверхностного фундамента происходит в следующей последовательности.Земляные работы: выработка котлована, выравнивание площадки, трамбовка поверхности, устройство подушки из слоя песка и слоя гравия.Укладка на подушку гидроизоляционных материалов.Заливка поверху гидроизоляции тонкого слоя бетона из подвижной смеси.Укладка арматуры.
Арматурный каркас делается из 2 арматурных сеток — верхней и нижней, которые жестко связывают вертикальными стержнями и проволокой.Между стержнями арматуры оставляют расстояние в 20—40 см. При этом получится в среднем 14 м арматурных стержней на 1 м2фундаментной плиты.Бетонирование котлована. Уплотнение бетона и уход за ним.На сооруженной плите может быть устроен монолитный ленточный фундамент, если требуется выполнить цоколь или подвал, но в этом случае плитный фундамент будет уже заглубленным, а не поверхностным.Возведение несущих стен здания (здесь как бы пропущен пункт об укладке перекрытия 1-го этажа, но этим перекрытием и станет плитный фундамент).В регионах с холодным климатом могут применяться морозоустойчивые плитные фундаменты с утолщенными ребрами.
При этом получится в среднем 14 м арматурных стержней на 1 м2фундаментной плиты.Бетонирование котлована. Уплотнение бетона и уход за ним.На сооруженной плите может быть устроен монолитный ленточный фундамент, если требуется выполнить цоколь или подвал, но в этом случае плитный фундамент будет уже заглубленным, а не поверхностным.Возведение несущих стен здания (здесь как бы пропущен пункт об укладке перекрытия 1-го этажа, но этим перекрытием и станет плитный фундамент).В регионах с холодным климатом могут применяться морозоустойчивые плитные фундаменты с утолщенными ребрами.
Толщина плиты берется та же (20—25 см), но края утолщаются, превращаясь в ребра. Ребра с некоторым шагом выполняют также по всей площади плиты.Обновлено 13.04.2012 07:36Дата: 21-12-2014Просмотров: 1007Рейтинг: 23Такое основание имеет несколько похожих названий — плавающий и незаглубленный. Чаще всего они исполнены в виде плит, хотя иногда можно встретить и ленточный поверхностный фундамент.Схема плавающего плитного фундамента. Иногда специалистами делается разграничение, при котором четко разделяется каждый из них:Плавающий — может быть изготовлен не только в форме плиты, но и иметь решетчатую структуру.
Иногда специалистами делается разграничение, при котором четко разделяется каждый из них:Плавающий — может быть изготовлен не только в форме плиты, но и иметь решетчатую структуру.
Назван он так, потому что перемещается вместе с грунтом, на котором устанавливается.Такой тип ни в коем случае нельзя устанавливать на сползающих грунтах, иначе однажды дом доползет до края участка, а потом начнет с него уползать, причем не метафорично, а в буквальном смысле (ползет грунт и тянет за собой дом).Незаглубленный — это общее название, которое определяет лишь степень заглубления, но не манеру установки (стационарный и плавающий). Иногда можно встретить не только плитный вариант или решетчатый, но ленточный и даже столбчатый, хоть и очень редко.Поверхностный — название для всех типов фундамента, которые не имеют заглубление, но при этом обязательным требованием является выполнение его в виде плиты. При этом совершенно не имеет значения, будет плита монолитной или же сборной из специальных блоков. Но если опустить формальные мелочи, можно выделить два варианта — ленточный фундамент и плитный, каждый из которых стоит отдельного внимания.Вернуться к оглавлениюСхема поверхностного фундамента.Для его возведения понадобятся:лопата штыковая;прут армирующий;аппарат сварочный;болгарка;рулетка;песок;раствор бетона М 250;дерево для опалубки;мусор строительный;вода и полиэтиленовая пленка.Такой вариант используется для небольших построек, которые служат временным нуждам — летняя кухня, баня, сарай и т.
Но если опустить формальные мелочи, можно выделить два варианта — ленточный фундамент и плитный, каждый из которых стоит отдельного внимания.Вернуться к оглавлениюСхема поверхностного фундамента.Для его возведения понадобятся:лопата штыковая;прут армирующий;аппарат сварочный;болгарка;рулетка;песок;раствор бетона М 250;дерево для опалубки;мусор строительный;вода и полиэтиленовая пленка.Такой вариант используется для небольших построек, которые служат временным нуждам — летняя кухня, баня, сарай и т.
п. Из жилых домов можно устанавливать только каркасные и другие аналоги легких деревянных построек. Важно, чтобы под будущей постройкой не оказалось случайно подземных вод, поскольку это неминуемо приведет к разрушению основания в первую же зиму в период вспучивания грунтов.Фундамент ленточный почти полностью исключает подземную часть работ, но при этом начать придется именно с них.Первым делом производится разметка будущего строения, которое крайне редко доходит до 9×10 м.
При копке следует держаться параметров траншеи — ширина и глубина равны 30 см. Вторым этапом изготавливается опалубка.Идеальным вариантом для нее станет ламинированная фанера, но можно применить ЛДСП или обрезную доску. Вся опалубка находится над поверхностью земли и имеет высоту 35 см. При больших показателях ленточный фундамент станет попутно и более хрупким.Армирующий каркас создается из ребристых прутов Ø12-15 мм.
Вторым этапом изготавливается опалубка.Идеальным вариантом для нее станет ламинированная фанера, но можно применить ЛДСП или обрезную доску. Вся опалубка находится над поверхностью земли и имеет высоту 35 см. При больших показателях ленточный фундамент станет попутно и более хрупким.Армирующий каркас создается из ребристых прутов Ø12-15 мм.
В горизонтальной обрешетке 3 уровня, всего таких обрешеток две.Все связи производятся при помощи сварочного аппарата и отрезов такого же прута. Важно, чтобы арматура не выходила за пределы бетона ни в одном месте. При установке под арматуру подкладывается строительный мусор (не пенного образования) для этой же цели.Схема мелкозаглубленного фундамента.На этом этапе желательно оборудовать все необходимые отводы (отверстия для труб).
Для этого к дереву монтируется отрезок трубы необходимого диаметра (обычно 7-8 см), причем делается это как можно более качественно: если он поменяет свое местоположение, можно считать, что отрез трубы стал частью арматуры, а отверстие делать придется перфоратором. После этого в опалубку заливается раствор бетона М 250 или большей марки. На застывание в среднем надо 3 недели при условии хорошей погоды. Если пойдет дождь, нужно укрыть ленточный фундамент полиэтиленом, но при этом всю первую неделю придется в любом случае поливать бетон водой, чтобы тот набрал максимальный запас прочности.Вернуться к оглавлениюДля него понадобятся инструменты и материалы, аналогичные указанным выше, а в дополнение — рубероид и подвижная смесь бетона.Плиты, как и ленты, используются для нетяжелых построек, но при этом они более податливы, а при передвижении качественная конструкция не пострадает.
После этого в опалубку заливается раствор бетона М 250 или большей марки. На застывание в среднем надо 3 недели при условии хорошей погоды. Если пойдет дождь, нужно укрыть ленточный фундамент полиэтиленом, но при этом всю первую неделю придется в любом случае поливать бетон водой, чтобы тот набрал максимальный запас прочности.Вернуться к оглавлениюДля него понадобятся инструменты и материалы, аналогичные указанным выше, а в дополнение — рубероид и подвижная смесь бетона.Плиты, как и ленты, используются для нетяжелых построек, но при этом они более податливы, а при передвижении качественная конструкция не пострадает.
Изготовление их более дорогое, но при вспучивании это дает свои результаты — фундамент может подниматься и опускаться, а на самом строении этот не отразится никоим образом.Для изготовления плитного поверхностного фундамента необходимо провести ряд следующих мероприятий:Земляные работы. Делается углубление в земле на 45 см, после чего грунт трамбуется.Засыпается песчаная подушка 5 см. Укладка гидроизоляции (рубероид).Поверх гидроизоляции заливается подвижная смесь бетона (толщина 10 см).Изготовление армирующего каркаса.Две горизонтальные обрешетки, у которых армирование не выходит за пределы бетона, связываются между собой отрезками прутьев. Все стыки делаются при помощи сварочного аппарата.
Укладка гидроизоляции (рубероид).Поверх гидроизоляции заливается подвижная смесь бетона (толщина 10 см).Изготовление армирующего каркаса.Две горизонтальные обрешетки, у которых армирование не выходит за пределы бетона, связываются между собой отрезками прутьев. Все стыки делаются при помощи сварочного аппарата.
Укладывать арматуруможно прямо на уже залитый раствор.Заливка котлована высокомарочным бетоном до краев (опалубка не нужна).Уход за раствором во время затвердевания аналогичен уходу за ленточным вариантом. Даже время высыхания примерно одинаковое, но иногда может доходить до 4-х недель. Никаких перекрытий между фундаментом и первым этажом делать не нужно, так как он по своей сути и является параллельно перекрытием.
Источники:
- domnasele.ru
- www.firmastr.ru
- moifundament.ru
Поверхностный фундамент | Инженерное решение
-
07.
 02.2012 05:08
02.2012 05:08
Данный тип фундамента называют также плитным, плавающим или незаглубленным. Поверхностный фундамент, как и все вышеописанные типы, широко применяется в индивидуальном жилом и дачном строительстве, обладая преимуществами по стоимости и простоте устройства.
Несмотря на то что все названия этого типа фундамента являются синонимами, по условиям возведения они отличны друг от друга.
Плитный фундамент может выполняться и заглубленным (с предварительной разработкой котлована), если необходимо устройство подвала или того требуют геологические условия. В индивидуальном строительстве заглубленный плитный фундамент применяется редко из-за своей дороговизны (поскольку на стоимость повлияют объемы земляных работ и большее по сравнению с ленточным фундаментом количество бетона и арматуры).
Поверхностный плитный фундамент, устроенный на подвижном пучинистом грунтовом основании, называется плавающим, поскольку перемещается вместе с грунтом без причинения вреда конструкциям дома. Плавающие поверхностные фундаменты могут выполняться как в форме сплошной плиты, так и в виде решетчатой структуры (монолитной или из сборных перекрестных железобетонных балок). В последнем случае необходима жесткая заделка узлов (мест стыков балок). Наибольшая жесткость плавающего фундамента достигается устройством плиты из монолитных перекрестных лент.
Плавающие поверхностные фундаменты могут выполняться как в форме сплошной плиты, так и в виде решетчатой структуры (монолитной или из сборных перекрестных железобетонных балок). В последнем случае необходима жесткая заделка узлов (мест стыков балок). Наибольшая жесткость плавающего фундамента достигается устройством плиты из монолитных перекрестных лент.
Плавающие фундаменты можно возводить на грунтах, подверженных сильной просадке, на насыпных грунтах (песчаная подушка, слежавшаяся свалка), на сильно пучинистых грунтах. Для плавающих фундаментов необходимо подбирать высокопрочный бетон и арматуру диаметром не менее 10 мм.
Незаглубленный фундамент сооружается не только в виде монолитной плиты, способен быть плавающим или стационарным (грунты не смещаются, поэтому и фундамент не двигается).
Поверхностный фундамент по смысловому значению почти полностью совпадает с незаглубленным, однако последний может быть также ленточным или столбчатым (хотя на практике такое встречается редко).
Поверхностный фундамент выполняется в виде сплошной плиты (чаще из монолитного железобетона или железобетонных блоков с обязательным армированием по всей площади и высоте). Причем земляные работы ограничиваются выравниванием площадки и разработкой неглубокого котлована.
Перед тем как выбрать данный тип фундамента, обратите внимание на следующие моменты.
1. Устраивать незаглубленный фундамент нужно только с учетом прочности основания (оно может быть пучинистым, но ни в коем случае не подверженным оползневым явлениям) и всех возможных деформаций. Причем надземные конструкции дома учитываются не только при расчете нагрузки на основание, но и при определении жесткости постройки: чем она выше, тем незначительнее будет деформироваться грунтовое основание и меньше риска при устройстве поверхностного фундамента.
2. Стоимость сооружения поверхностного плитного фундамента повышается за счет большого расхода материалов (ведь фундамент устраивается под всей площадью здания), поэтому целесообразно применять этот тип фундамента при возведении простых в плане зданий. Однако при составлении сметы надо учесть и небольшую стоимость земляных работ, и общий объем фундамента (а не только его площадь).
Однако при составлении сметы надо учесть и небольшую стоимость земляных работ, и общий объем фундамента (а не только его площадь).
3. Из-за большого расхода материалов плитный фундамент чаще применяется в небольших зданиях, где сама плита фундамента способна служить полом (гараж, баня). Но если сделать поверхностный фундамент не в виде сплошной плиты, а в виде ребристой монолитной плиты или армированных перекрестных лент, то такой тип фундамента будет хорошим вариантом и для жилых домов высокого класса.
Плитный фундамент в здании с подвалом может обойтись дороже, чем ленточный, поэтому часто застройщики возводят ленточный фундамент, а пол цокольного или подвального этажа устраивают отдельно. Однако подобная конструкция в отличие от плитного фундамента не обеспечивает надежную гидроизоляцию и больше подвержена просадкам.
4. Данный тип фундамента обладает существенным достоинством: если конструкция его жесткая (монолитная плита), то при выполнении такого фундамента под всей площадью здания на его конструкции не будут существенно влиять перемещения грунта (как сезонные, так и любые другие). Фундамент станет двигаться вместе с грунтовым основанием (отсюда и название «плавающий»), поэтому конструкции не будут деформироваться и трескаться. Но именно поэтому грунтовое основание не должно быть очень подвижным (иначе дом может в буквальном смысле слова «уползти» от вас). По отношению же к пучинистости (воздействию замораживания, оттаивания и просадки грунта) такой фундамент вполне устойчив.
Фундамент станет двигаться вместе с грунтовым основанием (отсюда и название «плавающий»), поэтому конструкции не будут деформироваться и трескаться. Но именно поэтому грунтовое основание не должно быть очень подвижным (иначе дом может в буквальном смысле слова «уползти» от вас). По отношению же к пучинистости (воздействию замораживания, оттаивания и просадки грунта) такой фундамент вполне устойчив.
5. Сооружение плитного поверхностного фундамента целесообразно на грунтовом основании с высокой степенью пучинистости и просадки (вертикальная подвижность грунта сильная, а горизонтальная — умеренная).
6. Если требуется высокий цоколь, лучше остановить выбор на ленточном фундаменте. Когда же достаточно невысокого цоколя, верх плитного фундамента способен послужить цокольным перекрытием.
От степени пучинистости грунта зависят толщина плиты основания и способ выполнения плитного фундамента:
— сильное пучение предполагает устройство фундамента из монолитного железобетона или готовых блоков, жестко скрепленных между собой;
— пучение средней силы позволяет возводить фундамент из монолитного железобетона или блоков, уложенных в перевязке на растворе;
— если пучение слабое, но все же присутствует, подойдут цементогрунт, бутобетон, бут, керамзитобетон.
Минимальная толщина плитного фундамента — 10 см. Чаще применяется плита толщиной 20—25 см, поскольку дальнейшее увеличение толщины приведет к росту нагрузок на слабый грунт, а применение минимальной толщины (10 см) не удобно для армирования и бетонирования (как следствие — понижение качества работ).
Для закладки плитного фундамента необходимы те же материалы и инструменты, что и для монолитного ленточного фундамента.
Требования к арматуре более высокие, чем в других типах фундамента. Она должна быть обязательно ребристого сечения, диаметром 12—16 мм.
Устройство поверхностного фундамента происходит в следующей последовательности.
- Земляные работы: выработка котлована, выравнивание площадки, трамбовка поверхности, устройство подушки из слоя песка и слоя гравия.
- Укладка на подушку гидроизоляционных материалов.
- Заливка поверху гидроизоляции тонкого слоя бетона из подвижной смеси.
- Укладка арматуры. Арматурный каркас делается из 2 арматурных сеток — верхней и нижней, которые жестко связывают вертикальными стержнями и проволокой.
 Между стержнями арматуры оставляют расстояние в 20—40 см. При этом получится в среднем 14 м арматурных стержней на 1 м2 фундаментной плиты.
Между стержнями арматуры оставляют расстояние в 20—40 см. При этом получится в среднем 14 м арматурных стержней на 1 м2 фундаментной плиты. - Бетонирование котлована. Уплотнение бетона и уход за ним.
- На сооруженной плите может быть устроен монолитный ленточный фундамент, если требуется выполнить цоколь или подвал, но в этом случае плитный фундамент будет уже заглубленным, а не поверхностным.
- Возведение несущих стен здания (здесь как бы пропущен пункт об укладке перекрытия 1-го этажа, но этим перекрытием и станет плитный фундамент).
В регионах с холодным климатом могут применяться морозоустойчивые плитные фундаменты с утолщенными ребрами. Толщина плиты берется та же (20—25 см), но края утолщаются, превращаясь в ребра. Ребра с некоторым шагом выполняют также по всей площади плиты.
Обновлено 13.04.2012 07:36
Виды поверхностных фундаментов
Варианты поверхностных фундаментов
Такое основание имеет несколько похожих названий – плавающий и незаглубленный. Чаще всего они исполнены в виде плит, хотя иногда можно встретить и ленточный поверхностный фундамент.
Чаще всего они исполнены в виде плит, хотя иногда можно встретить и ленточный поверхностный фундамент.
Схема плавающего плитного фундамента.
Иногда специалистами делается разграничение, при котором четко разделяется каждый из них:
- Плавающий – может быть изготовлен не только в форме плиты, но и иметь решетчатую структуру. Назван он так, потому что перемещается вместе с грунтом, на котором устанавливается. Такой тип ни в коем случае нельзя устанавливать на сползающих грунтах, иначе однажды дом доползет до края участка, а потом начнет с него уползать, причем не метафорично, а в буквальном смысле (ползет грунт и тянет за собой дом).
- Незаглубленный – это общее название, которое определяет лишь степень заглубления, но не манеру установки (стационарный и плавающий). Иногда можно встретить не только плитный вариант или решетчатый, но ленточный и даже столбчатый, хоть и очень редко.
- Поверхностный – название для всех типов фундамента, которые не имеют заглубление, но при этом обязательным требованием является выполнение его в виде плиты.
 При этом совершенно не имеет значения, будет плита монолитной или же сборной из специальных блоков.
При этом совершенно не имеет значения, будет плита монолитной или же сборной из специальных блоков.
Но если опустить формальные мелочи, можно выделить два варианта – ленточный фундамент и плитный, каждый из которых стоит отдельного внимания.
Вернуться к оглавлению
Поверхностный ленточный фундамент
Схема поверхностного фундамента.
Для его возведения понадобятся:
- лопата штыковая;
- прут армирующий;
- аппарат сварочный;
- болгарка;
- рулетка;
- песок;
- раствор бетона М 250;
- дерево для опалубки;
- мусор строительный;
- вода и полиэтиленовая пленка.
Такой вариант используется для небольших построек, которые служат временным нуждам – летняя кухня, баня, сарай и т.п. Из жилых домов можно устанавливать только каркасные и другие аналоги легких деревянных построек. Важно, чтобы под будущей постройкой не оказалось случайно подземных вод, поскольку это неминуемо приведет к разрушению основания в первую же зиму в период вспучивания грунтов.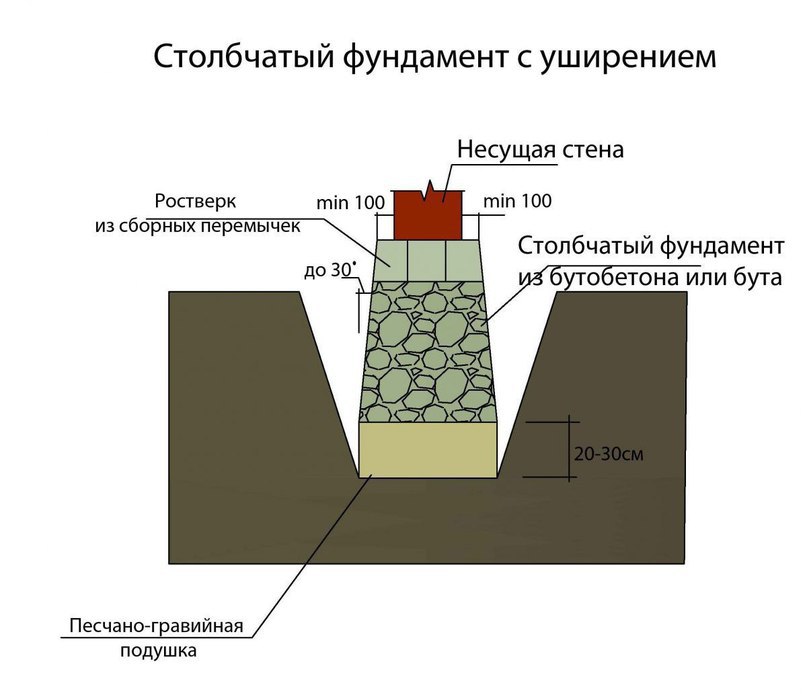
Фундамент ленточный почти полностью исключает подземную часть работ, но при этом начать придется именно с них.
Первым делом производится разметка будущего строения, которое крайне редко доходит до 9×10 м. При копке следует держаться параметров траншеи – ширина и глубина равны 30 см.
Вторым этапом изготавливается опалубка. Идеальным вариантом для нее станет ламинированная фанера, но можно применить ЛДСП или обрезную доску. Вся опалубка находится над поверхностью земли и имеет высоту 35 см. При больших показателях ленточный фундамент станет попутно и более хрупким.
Армирующий каркас создается из ребристых прутов Ø12-15 мм. В горизонтальной обрешетке 3 уровня, всего таких обрешеток две. Все связи производятся при помощи сварочного аппарата и отрезов такого же прута. Важно, чтобы арматура не выходила за пределы бетона ни в одном месте. При установке под арматуру подкладывается строительный мусор (не пенного образования) для этой же цели.
Схема мелкозаглубленного фундамента.
На этом этапе желательно оборудовать все необходимые отводы (отверстия для труб). Для этого к дереву монтируется отрезок трубы необходимого диаметра (обычно 7-8 см), причем делается это как можно более качественно: если он поменяет свое местоположение, можно считать, что отрез трубы стал частью арматуры, а отверстие делать придется перфоратором.
После этого в опалубку заливается раствор бетона М 250 или большей марки. На застывание в среднем надо 3 недели при условии хорошей погоды. Если пойдет дождь, нужно укрыть ленточный фундамент полиэтиленом, но при этом всю первую неделю придется в любом случае поливать бетон водой, чтобы тот набрал максимальный запас прочности.
Вернуться к оглавлению
Плитный поверхностный вариант основания
Для него понадобятся инструменты и материалы, аналогичные указанным выше, а в дополнение – рубероид и подвижная смесь бетона.
Плиты, как и ленты, используются для нетяжелых построек, но при этом они более податливы, а при передвижении качественная конструкция не пострадает. Изготовление их более дорогое, но при вспучивании это дает свои результаты – фундамент может подниматься и опускаться, а на самом строении этот не отразится никоим образом.
Изготовление их более дорогое, но при вспучивании это дает свои результаты – фундамент может подниматься и опускаться, а на самом строении этот не отразится никоим образом.
Для изготовления плитного поверхностного фундамента необходимо провести ряд следующих мероприятий:
- Земляные работы. Делается углубление в земле на 45 см, после чего грунт трамбуется.
- Засыпается песчаная подушка 5 см.
- Укладка гидроизоляции (рубероид).
- Поверх гидроизоляции заливается подвижная смесь бетона (толщина 10 см).
- Изготовление армирующего каркаса. Две горизонтальные обрешетки, у которых армирование не выходит за пределы бетона, связываются между собой отрезками прутьев. Все стыки делаются при помощи сварочного аппарата. Укладывать арматуру можно прямо на уже залитый раствор.
- Заливка котлована высокомарочным бетоном до краев (опалубка не нужна).
Уход за раствором во время затвердевания аналогичен уходу за ленточным вариантом. Даже время высыхания примерно одинаковое, но иногда может доходить до 4-х недель. Никаких перекрытий между фундаментом и первым этажом делать не нужно, так как он по своей сути и является параллельно перекрытием.
Никаких перекрытий между фундаментом и первым этажом делать не нужно, так как он по своей сути и является параллельно перекрытием.
типы опор, инструкция по возведению, советы
Содержание
- Виды опорно-столбчатых оснований
- Опоры из блоков
- Этапы возведения опорно-столбчатого основания
- Опоры из бетонной смеси
- Опоры из кирпича
- Опорно-столбчатый фундамент поверхностного типа
- Как избежать типичных ошибок
Наиболее часто данный тип основания применяется для небольших легких построек. Основным преимуществом является не только то, что опорно-столбчатый фундамент своими руками построить достаточно легко, но и то, что его стоимость будет гораздо дешевле, нежели возведение ленточного основания даже мелкого заложения.
к содержанию ↑
Виды опорно-столбчатых оснований
Прежде чем говорить о популярности и целесообразности применения данного типа основания для той или иной постройки вначале следует разобраться, а что такое опорно-столбчатый фундамент? Конструкция представляет собой заглубленные на определенную глубину опоры, которые впоследствии соединяются между собой ростверком.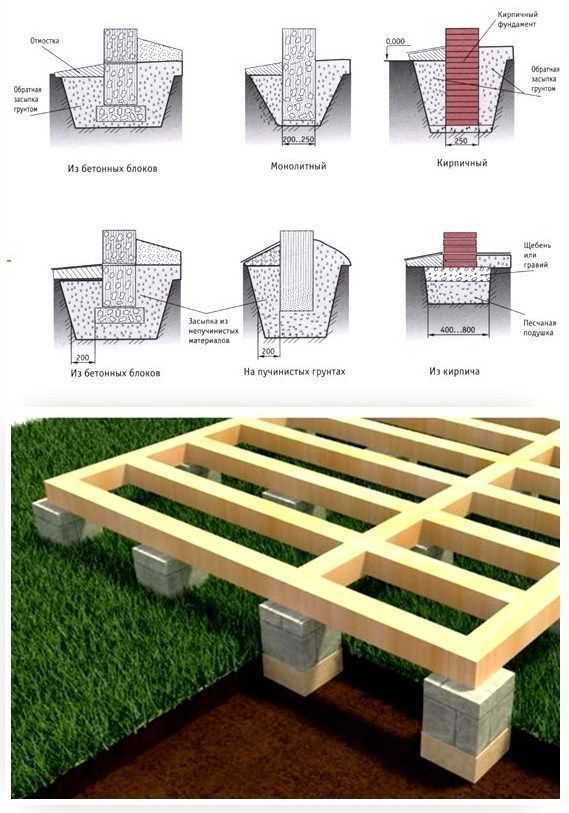
Данный тип основания можно устраивать на любых грунтах, в том числе и на суглинистых, склонных к пучению. Несмотря на легкость возведения и меньший объем работ по сравнению с монолитной конструкцией. Такое основание обеспечивает высокую несущую прочность конструкции.
Типы опорно-столбчатого фундамента зависят исключительно от материала, который будет использован для него:
- бетонные;
- блочные;
- бутовые;
- кирпичные;
- бутобетонные.
В редких случаях в качестве опор при этом основании может быть использована древесина, но такой фундамент подойдет только для хозяйственных построек ввиду своей малой несущей способности и небольшой долговременности.
В настоящее время большую популярность приобрели столбчатые фундаменты в качестве опор, для которых используются бетонные, кирпичные или блочные опоры.
Так, кирпичные опоры вполне могут обеспечить высокую прочность и долговременность эксплуатации, но они имеют один большой минус.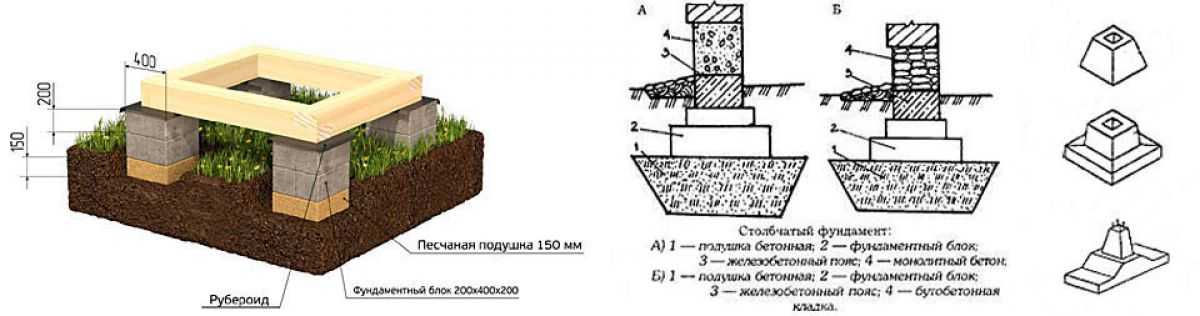 Фундамент из такого материала процесс достаточно трудоемкий. Не каждый вид кирпича подойдет для этого процесса.
Фундамент из такого материала процесс достаточно трудоемкий. Не каждый вид кирпича подойдет для этого процесса.
к содержанию ↑
Опоры из блоков
Опорно-столбчатый фундамент из блоков сегодня завоевал наибольшую популярность. Это объясняется более короткими сроками возведения основы под постройку (если сравнивать с другими материалами для опорных столбов), экономичностью и высокой прочностью конструкции в результате.
У опорных столбов из блоков есть существенный недостаток. Под них необходимо сделать хорошую гидроизоляцию из бетонной смеси или положить сверху песчано-гравийной подошвы рубероид. Это обусловлено тем, что блоки ввиду своей пористой структуры впитывают воду, которая приводит к их разрушению.
Этот недостаток легко устраняется при устройстве гидроизоляции не только под опорами. После возведения столбы обмазывают битумной мастикой в 2 или 3 слоя и н покрывают сверху геотекстилем.
Специалисты крайне не рекомендуют использовать такое основание на проблемных, пучинистых грунтах, а также на почвах с высокой влажностью и сейсмически не устойчивых. На проблемных грунтах целесообразнее использовать опоры, выполненные из бетонной смеси.
На проблемных грунтах целесообразнее использовать опоры, выполненные из бетонной смеси.
к содержанию ↑
Этапы возведения опорно-столбчатого основания
- Первоначально, как и при возведении любого основания необходимо выполнить подготовительные работы. Они заключаются в проведении изысканий для определения свойств почвы на строительном участке, расчетах конструкции и материла. После этого можно переходить к разметочным работам на участке.
- Далее на участке выкапывают или пробуривают скважины для будущих опор. При этом следует учитывать, что столбы должны быть установлены не только в крайних угловых точках, но и под продольными стенами, а также в местах пересечения внутренних стен будущей постройки.
- Размеры скважины должны быть больше размеров будущей опоры, поскольку необходимо учитывать еще толщину опалубки и толщину слоя гидроизоляции.
- Для того чтобы обеспечить столбам хорошую устойчивость и минимизировать возможную просадку, на дне выкопанных ям почву тщательно трамбуют.
 После этого устраивают гидроизоляционную подошву. Для этого используют смесь песка и гравия. Слой такой подошвы должен быть не менее 10 см. Отсыпав материал, его увлажняют водой и тщательно утрамбовывают.
После этого устраивают гидроизоляционную подошву. Для этого используют смесь песка и гравия. Слой такой подошвы должен быть не менее 10 см. Отсыпав материал, его увлажняют водой и тщательно утрамбовывают. - Далее, в зависимости от материала выполняют непосредственно возведение столбов.
к содержанию ↑
Опоры из бетонной смеси
Если в качестве основного несущего элемента будут использоваться бетонные столбы, то выполняют следующие операции.
- Изготавливают опалубку. Для этого вполне подойдут доски небольшой толщины – 4 см. в некоторых случаях можно сделать несъемную опалубку. Для этого целесообразно использовать трубы из железа или асбестоцемента, диаметром от 10 см.
- Бетонные столбы обязательно должны быть армированы. В противном случае от неукрепленного арматурой основания не будет никакого толка, а постройка вряд ли простоит без деформаций много лет.
- Прутки арматуры диаметром от 2 до 14 мм скрепляют между собой вязальной проволокой, но так чтобы вертикальный размер прутков был больше глубины опоры.
 Это необходимо, чтобы при дальнейших работах приварить к концам вертикальных прутков металлический каркас ростверка. Достаточно будет оставить 20 см.
Это необходимо, чтобы при дальнейших работах приварить к концам вертикальных прутков металлический каркас ростверка. Достаточно будет оставить 20 см. - Когда арматурный каркас подготовлен и опущен в яму, можно заливать бетонный раствор. Во время заливки необходимо тщательно штыковать его, чтобы максимально снизить образование воздушных пустот, приводящих к понижению прочности столбов.
- Когда опоры готовы, приступают к строительству ростверка. Этот элемент предназначен для связывания отдельных опорных элементов в одну конструкцию, которая позволит равномерно распределить нагрузки по всему основанию.
- Сначала к вертикальным пруткам опор приваривается арматурный каркас. Затем устанавливается опалубка, и бетонная смесь заливается в форму.
к содержанию ↑
Опоры из кирпича
Когда точечный опорно-столбчатый фундамент возводится с опорамииз кирпича, необходимо позаботиться о хорошей гидроизоляции. Для этого на песчано-гравийную подошву укладывают рубероид и выкладывают кирпичи.
Опорные столбы из кирпича необходимо тщательно заизолировать от воды и влажного грунта. Для этого поверхность опоры нужно тщательно заштукатурить. Желательно в раствор цемента добавить немного жидкого стекла.
После того как штукатурка станет прочной, ее поверхность желательно обработать грунтовкой, имеющей антибактериальные свойства. Более простым вариантом гидроизоляции является битумная мастика. Ее наносят в 2-3 слоя, сверху необходимо наклеить геотекстиль, Он необходим для защиты опорных столбов от возможных механических повреждений, вызванных движением грунта.
к содержанию ↑
Опорно-столбчатый фундамент поверхностного типа
Отдельно следует рассмотреть и поверхностный опорно-столбчатый фундамент. Его устраивают для небольших построек, возводимых преимущественно из дерева или бруса: бани, дачные домики и так далее.
Опоры необходимо устанавливать под углами здания, несущими конструкциями. Этот тип основания целесообразно использовать только на устойчивых, плотных грунтах, поскольку глубина заложения опорных столбов небольшая. Для суглинистых почв, грунтов с повышенной влажностью, почв с близко расположенными к поверхности грунтовыми водами, а также на участках с большими перепадами высот такой фундамент устраивать нельзя, независимо от общей массы будущей конструкции.
Для суглинистых почв, грунтов с повышенной влажностью, почв с близко расположенными к поверхности грунтовыми водами, а также на участках с большими перепадами высот такой фундамент устраивать нельзя, независимо от общей массы будущей конструкции.
к содержанию ↑
Как избежать типичных ошибок
Как сделать опорно-столбчатый фундамент вы знаете, но самое главное это постараться при его возведении, избежать распространенных ошибок. Зачастую, возводя основание своими силами, многие путают столбчатый и свайный фундамент и устанавливают опорные столбы по той же технологии, что и сваи. То есть в заранее пробуренные скважины забивают столбы.
На самом деле, эти два типа основания совершенно разные. Опорно-столбчатый фундамент подходит только для легких конструкций: бани, одноэтажные, небольшие по площади каркасные дома или строения из бруса.
Свайный же фундамент выдерживает достаточно большой вес постройки. Сваи, используемые в качестве столбов нельзя устанавливать в заранее пробуренные скважины большего диаметра, поскольку прочность их установки как раз и зависит от того, что их заглубляют в землю при помощи приваренных к ним лопастей.
В заключение можно сказать, что для небольших построек столбчатое основание является самым оптимальным и недорогим вариантом. Но если вы собираетесь установить такой тип фундамента под дом, то обязательно учтите, что это основание нуждается в дополнительной теплоизоляции, поскольку подвала оно не имеет. И если вы не хотите платить зимой за уходящее сквозь пол тепло об этих работах необходимо позаботиться еще на этапе подготовительных работ.
Расширенное проектирование поверхностных фундаментов
Модуль Расширенное проектирование поверхностных фундаментов дополняет модули Фундаменты и Оголовки свай , когда они используются в CYPECAD. Позволяет проектировать фундаменты с фундаментами и ростверками с особыми пересечениями элементов (пересечения ленточных и анкерных балок), геометрической обрезкой фундаментов и приложением линейных, точечных и поверхностных нагрузок на фундаменты, ростверки и ленточные и анкерные балки. В случае CYPE 3D этот модуль позволяет выполнять обрезку только геометрии фундамента.
Позволяет проектировать фундаменты с фундаментами и ростверками с особыми пересечениями элементов (пересечения ленточных и анкерных балок), геометрической обрезкой фундаментов и приложением линейных, точечных и поверхностных нагрузок на фундаменты, ростверки и ленточные и анкерные балки. В случае CYPE 3D этот модуль позволяет выполнять обрезку только геометрии фундамента.
Полигональные ограничения для фундаментов (в CYPECAD и CYPE 3D)
С помощью модуля «Расширенное проектирование поверхностных фундаментов» в CYPECAD или CYPE 3D пользователь может определить пределы или границы, которые не могут быть нарушены железобетонными или монолитными фундаментами. Примеры этих пределов или границ могут включать пределы собственности или зоны, зарезервированные для других целей, таких как месторождения, лифтовые шахты и т. д. Эти пределы также могут быть определены для программы для определения неправильных многоугольных фундаментов по любой другой причине, независимо от того, существуют ли реальные физические ограничения. пределы.
пределы.
Пользователь должен ввести ломаные линии, чтобы задать пределы. Когда программа проектирует фундамент, она автоматически обрезает фундаменты, которые выходят за установленные пределы или выходят за их пределы. Часть фундамента, в которой начинается колонна или остаются колонны, сохраняется и, следовательно, учитывается при анализе. При проектировании фундаментов программа не допускает превышения установленных пределов. На конструкцию фундамента может влиять более чем одно ограничение.
Ограничения вводятся в CYPECAD с помощью опции Ограничения для многоугольных фундаментов , расположенной в меню «Фундаменты» на вкладке «Определение колонны» или на вкладке «Результаты». В CYPE 3D эта опция доступна на вкладке «Основы» > «Ограничения» > «Создать». Ограничения можно ввести вручную, определив каждое ребро в обеих программах или привязав к шаблону DXF/DWG. В CYPECAD ограничения, введенные в группу, влияют только на фундаменты этой группы, в то время как в CYPE 3D ограничения влияют на все фундаменты, независимо от их высоты.
После того, как полилинии, определяющие пределы, введены, пользователь может удалить или переместить их, вставить новую вершину или удалить участок полилинии.
Эти полилинии имеют определенные логические ограничения относительно их положения:
- Они не могут пересекать колонну.
- Они должны вырезать контуры фундаментов до и после их проектирования.
- Фундаменты с несколькими колоннами нельзя обрезать таким образом, чтобы одна колонна оставалась изолированной от остальных колонн или колонна начинала совместно использовать фундамент.
- Определенные ограничения не действуют на ленточные и анкерные балки, наголовники свай, конические или ленточные фундаменты. Они воздействуют только на железобетонные и массивные бетонные основания постоянной глубины. Тем не менее, CYPECAD позволяет своим пользователям определять комбинированное основание постоянной глубины, содержащее несколько опор, которые также могут включать стену или стену сдвига (определяемую с внешней фиксацией и без основания), и в этом случае пределы будут влиять на основание.
 .
.
Если ломаная линия не соответствует одному из этих условий, программа не будет учитывать ограничение, созданное ломаной линией. В этом случае объяснение, почему не удалось адаптировать контур фундамента к введенным ограничениям, приводится в акте проверки каждого затронутого фундамента.
Возможные причины:
- Введенное ограничение прорезает столбец.
- Введенный предел пересекает фундамент, но не разделяет его.
- Предел полностью находится внутри фундамента.
- Поскольку в фундаменте нет зачатков арматуры, эффективный контур не может быть рассчитан.
- Предел разделяет фундамент без начала армирования, лежащего в одной части фундамента.
- На конические фундаменты введенные ограничения не распространяются.
Отчеты о проверке фундамента можно визуализировать с помощью различных опций:
В CYPECAD:
- Определение балки или вкладка Результаты > Фундамент > Код ошибки проверки > Выбрать фундамент
- Вкладка Определение балки или Результаты > Фундамент > Элементы фундамента > значок «Редактировать» > «Выбрать основание» > кнопка «Проверка кода» или кнопка «Дизайн».

В CYPE 3D :
- Вкладка «Фундамент» > «Анализ» > «Проверить» > «Выбрать фундамент»
- Вкладка «Фундамент» > «Элементы фундамента» > «Выбрать фундамент» > кнопка «Проверить» или кнопка «Проект».
Пересечения ленточных и анкерных балок (только для CYPECAD)
Используя модуль Расширенное проектирование поверхностных фундаментов , CYPECAD может проектировать фундаменты, состоящие из фундаментов или оголовков свай с пересекающимися лентовидными и анкерными балками.
Нагрузки, прикладываемые к фундаментам, оголовкам свай, лямкам и анкерным балкам (только для CYPECAD)
Используя модуль Расширенное проектирование поверхностных фундаментов , CYPECAD позволяет вводить точечные, линейные и поверхностные нагрузки на фундаменты, оголовки свай, ленточные и анкерные балки и учитывать их при проектировании. Используя этот модуль, CYPECAD также распознает нагрузки, передаваемые опорами лестницы, опирающимися на вышеупомянутые элементы фундамента.
CYPECAD исходит из того, что грунт под ремнями и стяжками не влияет на равновесие или распределение сил, которым они подвергаются. Следовательно, ленточные и анкерные балки также рассчитаны на поддержку любых нагрузок, которые могут воздействовать на них, и будут передавать усилия на фундаменты или оголовки свай на их концах или на другие ленточные и анкерные балки, которые они могут пересекать.
Другие функции
Для доступа к дополнительным функциям, предлагаемым программой, существует несколько модулей, которые можно найти на веб-страницах «Модули CYPECAD» и «3D-модули CYPE».
Дополнительная информация
Учебные ресурсы
Техническая поддержка
Купить сейчас
9 Деревья, которые могут повредить ваш фундамент (и как исправить)
Ничто не сравнится с походом в лес или прогулкой так же хорош, как и красивый пейзаж, усеянный высокими деревьями. Деревья дают нам тень, кислород, очищение воздуха, и на них просто приятно смотреть, особенно осенью! Но чего мы можем не осознавать, так это того, что большая часть дерева лежит под землей, а под почвой скрывается проблема, ожидающая своего решения.
Корни деревьев могут повредить фундамент зданий, а также тротуары и тротуары. Наиболее распространенными деревьями, которые повреждают фундамент здания, являются остролистный клен, серебристый клен, дуб, ясень, тополь, грецкий орех, тополь и платан. Эти деревья имеют высокий потенциал для нанесения ущерба под землей.
Есть несколько вещей, которые делают ваш фундамент более восприимчивым к повреждению корней. Но будьте уверены, есть способы смягчить и даже предотвратить приближение корней к вашему фундаменту.
Просто добавим: когда вы совершаете покупки по ссылкам из Tree Journey, мы можем получать партнерские комиссионные, если вы совершаете покупку. Как партнер Amazon, мы зарабатываем на соответствующих покупках.
Могут ли деревья влиять на фундамент дома?
Прежде чем говорить о конкретных деревьях, которые могут нанести урон, давайте обсудим сам ущерб. Как именно деревья влияют на фундамент вашего дома?
По данным Департамента общественных работ Вайоминга, до 90% корней деревьев находятся в верхних 12 дюймах почвы. Фундаменты имеют глубину от нескольких дюймов ниже поверхности до нескольких футов.
Излишне говорить, что ваш фонд находится прямо в целевой зоне цепляющихся корней.
Существует два основных способа повреждения фундамента корнями. Они либо поднимут ваше здание, вызывая трещины, либо сожмут почву, что приведет к оседанию фундамента.
Деревья могут приподнять фундамент дома
Первый способ, которым корни могут повредить фундамент, — это слегка приподнять дом. Это оказывает давление на фундамент, который может прогнуться и треснуть.
Корни на здоровом дереве растут ежегодно, как и ствол. Университет Пердью утверждает, что обычно, когда корень сталкивается с непроницаемым объектом (вашим основанием), он стремится пройти вокруг вместо под .
Это особенно верно, потому что почва под вашим фундаментом, вероятно, уплотнена из-за первоначальной конструкции и высохла из-за покрытия поверх почвы.
Тем не менее, некоторые корни упрямы и полны решимости уйти под ваш фундамент или через уже существующие трещины.
По мере того, как корни продолжают расти, они будут занимать все больше и больше места под вашим домом, буквально на мельчайшие части поднимая здание.
Это явление встречается гораздо реже, чем другие корневые проблемы. У вас больше шансов найти корни, растущие вдоль фундамента, чем под ним.
Деревья могут вызвать усадку почвы рядом с фундаментом
Более распространенный способ повреждения фундамента корнями — усадка почвы. Итак, как именно корни сокращают почву?
Корни будут искать наиболее выгодный путь для роста. Это означает много влаги, рыхлую почву и много питательных веществ . Хотя это обычно не вызывает проблем, корни, которые проникают в глинистую почву, могут значительно изменить высоту почвы, когда они проходят.
Глинистые почвы обладают особым свойством: они могут удерживать тонну влаги. Но когда эта влага исчезает, глинистые почвы имеют тенденцию усыхать, и, , как вы уже догадались, , сжимайся!
Итак, когда корни всасывают всю воду, которую они могут получить, они на самом деле уплотняют почву вокруг себя. Это, в свою очередь, приводит к оседанию фундамента. В процессе осадки в фундаменте могут образоваться трещины.
Наиболее распространенные деревья, которые наносят ущерб вашему фундаменту
Независимо от того, строите ли вы новый дом или покупаете уже существующий, вам необходимо внимательно изучить ландшафт. Какие деревья вы видите?
Если вы не уверены, вы всегда можете воспользоваться справочником по деревьям Национальной федерации дикой природы, который поможет вам понять, на что вы смотрите!
Некоторые деревья более подвержены повреждению фундамента, чем другие. Согласно Мичиганского государственного университета , деревья, которые быстро растут над землей, часто так же быстро растут под землей.
Это деревья, на которые стоит обратить внимание, потому что их корни будут далеко и быстро достигать. Давайте погрузимся в это!
Дубы повреждают фундамент
Дубы не так распространены в ландшафте, как другие деревья, но, несмотря на это, они, как правило, наносят наибольший ущерб домам.
Существует множество различных видов дубов, около 90 только в Соединенных Штатах. Их обычно считают медленнорастущими, но они массивные , многие виды способны достигать высоты 100 футов и более.
Эти огромные дубовые организмы удерживаются на месте глубоким стержневым корнем, который прорастает прямо под стволом. По мере роста дерева боковые корни закрепляются и начинают извиваться от дерева во всех направлениях. Эти корни могут достигать трехкратной высоты дерева.
Помимо того, что некоторые виды дуба представляют опасность для вашего фундамента, они склонны к падению сучьев. Если у вас во дворе растет дуб, а сук висит над вашим домом, лучше удалить его, прежде чем он решит упасть сам.
Если вам нужно правильное удобрение для дуба, мы написали руководство по лучшим удобрениям для вашего дуба!
Корни ясеня вызывают повреждение фундамента
Ясени часто путают с некоторыми другими проблемными деревьями, такими как тополя, гикори и бузина.
Самая большая проблема с корнями ясеня заключается в том, что они неглубокие. После формирования глубокого стержневого корня формируются боковые корни, как большие, так и малые. По данным Лесной службы Министерства сельского хозяйства США, большие корни ясеня в равной степени можно найти в горизонте A почвы (поверхность почвы), а также в горизонте B (недра).
Чтобы узнать ясень, посмотрите на веточки! Они будут располагаться напротив друг друга на ветвях, а не чередоваться, как у большинства других лиственных деревьев.
Тополь повреждает фундамент
Третье дерево в нашем списке — тополь. Их часто путают с трясущимися осинами или березами.
Корни тополя обычно представляют собой смесь мелких и глубоких корней. Они плохо себя чувствуют в уплотненной почве, например, в результате строительства в городских и пригородных районах. Вместо этого они найдут небольшие карманы рыхлой почвы, чтобы прорасти свои корни.
Это делает старые фундаменты особенно восприимчивыми к корням тополя. Если у вас есть небольшая трещина или несовершенство в фундаменте, корень тополя найдет ее и войдет внутрь, расширив трещину.
Корни норвежского и серебристого клена повреждают фундамент
В отличие от дуба, ясеня и тополя, не все клены вредны для вашего фундамента. Два из них, которые вызывают наибольшую озабоченность, — это остролистный и серебристый клены.
И остролистный, и серебристый клены быстро растут. Корневая система остролистного клена настолько мелкая, что может конкурировать с вашей газонной травой за питательные вещества! Серебряные клены известны своей массивной и быстрорастущей корневой системой.
Если вы сажаете клен серебристый, рекомендуется сажать его не менее чем в 10 футах от фундамента. Больше, если сможешь.
Клен остролистный – одно из немногих известных деревьев, способных поднимать фундамент, а не повреждать его из-за усадки глинистой почвы.
Если вам нужно хорошее удобрение для клена, ознакомьтесь с нашим руководством по лучшим удобрениям для клена здесь!
Деревья черной акации быстро растут, и их корни могут повредить фундамент
Robinia pseudoacacia , черная акация, быстрорастущее дерево, произрастающее в восточной части США. Это дерево очень трудно контролировать после того, как оно укоренится, что делает его возможным проблемным деревом для домовладельцев.
Черная акация размножается так же, как и осина, путем создания клонов. Это делается с помощью процесса, называемого присасыванием , при котором новые побеги образуются из отмирающих корней или старых пней.
Помимо сложности управления этими выгодными деревьями, они также имеют очень широкую мочковатую корневую систему. Это может быть проблематично для фондов, особенно потому, что корни быстро растут.
На самом деле, лучше даже не смотреть на акацию…
Боксерские деревья могут поднять ваш фундамент
Боксельдеры не очень желательны во дворах, улицах или городских районах. Они привлекают клопов-боксеров, недолговечны и имеют хрупкую древесину, склонную к падению.
Однако, по данным Лесной службы США, они могут расти в самых разных условиях и появляться то тут, то там сами по себе во многих областях.
Подобно остролистному клену, самшиты имеют очень неглубокую корневую систему, которая может поднимать тротуары и фундаменты, а не уплотнять почву и вызывать ее оседание.
Не подходит для посадки рядом с домом.
Корни елей могут повредить фундамент
Два вида елей могут повредить фундамент. Ель плакучая Брюера и ель европейская. Кажется, что норвежские виды просто запрещены, верно?
По данным 2003 г. , исследование , корни ели обыкновенной предпочитают почвы, богатые гумусом, которые обычно находятся у поверхности в горизонте О.
Хотя у ели плакучей не такие мелкие корни, как у ели европейской, она все же цепляется за поверхность почвы, а несколько вертикальных корней проникают глубже, удерживая дерево на месте.
В общем, чем глубже уходят корни (и чем шире), тем выше вероятность возникновения проблем.
Другие деревья, которые повреждают фундамент
Несмотря на то, что список деревьев, которые могут нанести ущерб, очень длинный, вышеперечисленные деревья с наибольшей вероятностью могут повредить ваш фундамент.
Here 8 other trees that can damage your foundation:
- Sweetgum
- Walnut
- Hickory
- Cottonwood
- Sycamore
- Loblolly Pine
- Crimean Сосна
- Тюльпанное дерево
Эти деревья вряд ли навредят вашему фундаменту, но обладают потенциалом из-за их быстрорастущих и неглубоких корней.
Как узнать, повреждают ли корни деревьев фундамент?
Нередки случаи, когда фундамент трескается не из-за корней деревьев. Почва естественным образом растет и сжимается в зависимости от погодных условий, здания оседают на недавно выкопанной почве, и даже небольшие толчки и землетрясения могут причинить ущерб, даже если вы их не чувствуете.
Тем не менее, если вы заметите небольшие трещины вокруг фундамента, вам нужно как можно быстрее узнать, что их вызывает, чтобы избежать дорогостоящего ущерба.
На самом деле есть только один способ определить, повреждают ли корни деревьев ваш фундамент, — это физически заглянуть под землю возле вашего фундамента и проверить наличие корней.
По данным Университета Флориды, вы можете сделать это, раскопав участок вокруг вашего фундамента и найдя корни. Это можно сделать легко, если вы проведете линию от ближайшего дерева к вашему дому и просто обкопаете эту область вашего фундамента.
Не обманывайтесь, если вы не найдете корней на прямой линии от дерева. Корни могут быть далеко уходящими, поэтому копайте по несколько футов в каждом направлении, чтобы убедиться, что вы видите весь масштаб корневой системы.
Как восстановить поврежденный фундамент из деревьев
Владение домом может быть замечательным опытом. Вы получаете свое собственное пространство, вам не нужно беспокоиться о выплатах арендодателям или иметь дело с хрупкими, тонкими стенами между вами и вашим соседом.
Но работы много!
Последнее, с чем вам хотелось бы иметь дело, это с повреждением фундамента вашего дома. Буквально! Трещины и трещины в вашем фундаменте и тротуаре могут быть головной болью, но есть способы исправить и предотвратить повреждение от корней деревьев.
Вода для вашего фонда
Не волнуйтесь, вы все правильно прочитали! Это может показаться странным, но полив области вокруг вашего фундамента может быстро решить вашу проблему.
Обычно это работает лучше всего, если ваш фундамент построен на глинистой почве. Деревья, как правило, скупы на воду, высасывая все, что могут получить, и почти ничего не оставляя вокруг себя.
Когда корни накапливают воду, глинистая почва высыхает, сморщивается и сморщивается. Полив области вокруг вашего фундамента раздует почву и, возможно, закроет трещины сами по себе.
Даже если повреждение вашего фундамента не вызвано скупыми корнями деревьев, вы можете использовать этот метод для ремонта вашего фундамента.
Сажать менее инвазивные виды
Это решение не работает, если в вашем дворе уже есть инвазивные деревья. Если вы не хотите тратить деньги на их вырубку и посадку новых деревьев, вы можете застрять на своих нынешних ландшафтных деревьях.
Однако, если вы думаете о посадке новых деревьев или только что построили дом и у вас есть возможность выбрать деревья для ландшафта, выберите породы деревьев, которые менее вредны для фундамента.
Деревья, не подверженные повреждению зданий, включают:
- Кипарисы: Существует несколько различных типов кипарисов, но большинство из них имеют глубокий стержневой корень, сопровождаемый боковыми корнями. Эти корни обычно не ломают поверхность и не вызывают проблем при строительстве фундамента.
- Фруктовые деревья: Деревья из рода Prunus довольно безопасно сажать вокруг домов. К ним относятся сливовые, вишневые, персиковые, абрикосовые и миндальные деревья.
- Сосны: За исключением крымской, лоблолли и швейцарской сосны, сосны — отличный выбор для посадки во дворе. Их корни имеют тенденцию расти вниз, а не наружу, защищая ваш фундамент от надоедливых корней.
Используйте барьер из корней деревьев, чтобы спасти свой фундамент
Корневые барьеры предназначены для направления корней в определенном направлении, например, в сторону от вашего фонда. Они работают, создавая непроницаемый барьер, через который корни не могут пробиться и должны обойти его.
Что-то вроде Барьер для корней деревьев DeepRoot может помочь отвести разрушительные корни от фундамента, мощеной дороги или тротуара. Самое замечательное в продукте DeepRoot то, что он поставляется в различных размерах, поэтому вы можете выбрать то, что лучше всего подходит для вашей ситуации.
Корневые барьеры особенно эффективны для неглубоких корневых систем, которыми обладает большинство деревьев, наносящих ущерб. Вы даже можете использовать их на неповрежденных корнях, таких как кусты, посаженные рядом с домом.
Докопаться до сути проблемы!
Деревья могут добавить много ценности вашему ландшафту, они обеспечивают убежище, пищу и тень множеству маленьких существ, за которыми может быть интересно наблюдать. Но мы не хотим, чтобы наши деревья причиняли вред нашим домам.
Такие деревья, как дубы, живут довольно долго. Дубы обычно имеют долгую жизнь. Таким образом, эти фундаментальные проблемы будут только расти!
Если вы заметили трещины или трещины в фундаменте, корни не всегда виноваты. Не забудьте проверить, какие деревья растут вокруг вашего дома, чтобы выяснить, есть ли среди них одно из самых опасных деревьев.
Напомним, деревья, которые с наибольшей вероятностью могут повредить ваш фундамент, включают:
- Oak
- Ash
- Poplar
- Silver Maple
- Norway Maple
- Black Locust
- Boxelder
- Norway Spruce
- Brewer’s Плакучая ель
Существует множество других потенциально опасных деревьев, но об этих 9 говорят чаще всего и они самые разрушительные.
Деревья с неглубокими корнями, которые не так далеко уходят, имеют свои проблемы. Эти деревья, как правило, подвержены повреждению ветром и могут сдуться легче, чем деревья с глубокими стержневыми корнями и далеко идущими боковыми корнями.
Принятие мер по предотвращению проникновения корней в фундамент до того, как у вас возникнет проблема, — лучший подход к решению проблем с инвазивными корнями. Но в целом деревья, как правило, оказывают более положительное влияние на наши дворы, чем отрицательное, особенно если вы знаете и готовы к блуждающим корням.
Ссылки:
Биддл, Г. (2001, 10-13 октября). Повреждение корней деревьев зданиям [Заседания Комитета по мелководью и свойствам грунта на конференции ASCE по гражданскому строительству]. Хьюстон, Техас. https://ascelibrary.org/doi/abs/10.1061/40592(270)1
Хитчмо, Дж., и Филдхаус, К. (2008). Руководство пользователя установки: Руководство по эффективной спецификации . Джон Уайли и сыновья.
Рэндруп, Т. Б., Макферсон, Э. Г., и Костелло, Л. Р. (2001 г., март). Проникновение корней деревьев в канализационные системы: обзор масштабов и затрат. Журнал инфраструктурных систем , 7 (1). https://ascelibrary.org/doi/abs/10.1061/(ASCE)1076-0342(2001)7:1(26)
Робертс, Дж., Джексон, Н., и Смит, М. (2006). Корни деревьев в искусственной среде . Канцелярия.
14.2 Первичные ткани животных – концепции биологии – 1-е канадское издание
Глава 14. Тело животного: основная форма и функция
Цели обучения
К концу этого раздела вы сможете:
- Опишите эпителиальные ткани
- Обсудите различные типы соединительной ткани у животных
- Опишите три типа мышечной ткани
- Опишите нервную ткань
Многоклеточные сложные животные имеют четыре основных типа тканей: эпителиальную, соединительную, мышечную и нервную. Напомним, что ткани представляют собой группы однотипных клеток, выполняющих родственные функции. Эти ткани объединяются, образуя органы, такие как кожа или почки, которые выполняют определенные специализированные функции в организме. Органы организованы в системы органов для выполнения функций; примеры включают систему кровообращения, состоящую из сердца и кровеносных сосудов, и пищеварительную систему, состоящую из нескольких органов, включая желудок, кишечник, печень и поджелудочную железу. Системы органов объединяются, чтобы создать целый организм.
Эпителиальные ткани
Эпителиальные ткани покрывают снаружи органы и структуры тела и выстилают просветы органов одним или несколькими слоями клеток. Типы эпителия классифицируются по форме присутствующих клеток и количеству слоев клеток. Эпителии, состоящие из одного слоя клеток, называются простыми эпителиями ; эпителиальная ткань, состоящая из нескольких слоев, называется многослойным эпителием . В табл. 14.2 приведены различные типы эпителиальных тканей.
| Форма ячейки | Описание | Местоположение |
|---|---|---|
| плоскоклеточный | плоская, неправильной круглой формы | простой: альвеолы легких, капилляры многослойные: кожа, рот, влагалище |
| прямоугольный | в форме куба, с центральным ядром | железы, почечные канальцы |
| столбчатый | высокое, узкое, ядро направлено к основанию высокое, узкое, ядро расположено вдоль клетки | простые: пищеварительный тракт псевдослоистые: дыхательные пути |
| переходной | круглый, простой, но выглядит многослойным | мочевой пузырь |
Плоскоклеточный эпителий
Плоскоклеточный эпителий обычно круглые, плоские клетки с небольшим центрально расположенным ядром. Очертания клеток слегка неровные, и клетки соединяются вместе, образуя покрытие или выстилку. Когда клетки располагаются в один слой (простой эпителий), они облегчают диффузию в ткани, например, в зоны газообмена в легких и обмен питательных веществ и отходов в кровеносных капиллярах.
Рис. 14.7 a иллюстрирует слой плоскоклеточных клеток, мембраны которых соединены вместе, образуя эпителий. Изображение На рис. 14.7 b показаны клетки плоского эпителия, расположенные в многослойных слоях, где необходима защита тела от внешнего истирания и повреждений. Это называется многослойным плоским эпителием и встречается в коже и тканях, выстилающих рот и влагалище.
Кубический эпителий
Клетки кубического эпителия , показанные на рис. 14.8, имеют форму куба с одним центральным ядром. Чаще всего они встречаются в виде одного слоя, представляющего собой простой эпителий в железистых тканях по всему телу, где они подготавливают и секретируют железистый материал. Они также обнаруживаются в стенках канальцев и в протоках почек и печени.
Столбчатый эпителий
Столбчатый эпителий Клетки больше в высоту, чем в ширину: они напоминают стопку столбцов в эпителиальном слое и чаще всего встречаются в однослойном расположении. Ядра столбчатых эпителиальных клеток в пищеварительном тракте кажутся выстроенными у основания клеток, как показано на рис. 14.9. Эти клетки поглощают материал из просвета пищеварительного тракта и подготавливают его к поступлению в организм через кровеносную и лимфатическую системы.
Рисунок 14.9. Простые столбчатые эпителиальные клетки поглощают материал из пищеварительного тракта. Бокаловидные клетки выделяют слизь в просвет пищеварительного тракта.Столбчатые эпителиальные клетки, выстилающие дыхательные пути, кажутся стратифицированными. Однако каждая клетка прикреплена к базовой мембране ткани и, следовательно, это простые ткани. Ядра расположены на разных уровнях в слое клеток, что создает впечатление наличия более одного слоя, как показано на рис. 14.10. это называется псевдомногослойный , цилиндрический эпителий. Это клеточное покрытие имеет реснички на апикальной или свободной поверхности клеток. Реснички усиливают движение слизистых и захваченных частиц из дыхательных путей, помогая защитить систему от инвазивных микроорганизмов и вредных материалов, которые вдыхаются в организм. Бокаловидные клетки вкраплены в некоторые ткани (например, в слизистую оболочку трахеи). Бокаловидные клетки содержат слизь, которая улавливает раздражители, которые в случае трахеи препятствуют попаданию этих раздражителей в легкие.
Рисунок 14.10. Псевдостратифицированный цилиндрический эпителий выстилает дыхательные пути. Они существуют в одном слое, но расположение ядер на разных уровнях создает впечатление, что существует более одного слоя. Бокаловидные клетки, расположенные между цилиндрическими эпителиальными клетками, выделяют слизь в дыхательные пути.Переходный эпителий
Переходные или уроэпителиальные клетки появляются только в мочевой системе, преимущественно в мочевом пузыре и мочеточнике. Эти клетки располагаются в виде стратифицированного слоя, но могут казаться нагроможденными друг на друга в расслабленном пустом мочевом пузыре, как показано на рис. 14.11. Когда мочевой пузырь наполняется, эпителиальный слой разворачивается и расширяется, удерживая введенный в него объем мочи. Когда мочевой пузырь наполняется, он расширяется, а слизистая оболочка становится тоньше. Другими словами, ткань переходит из толстой в тонкую.
Рисунок 14.11. Переходный эпителий мочевого пузыря претерпевает изменения толщины в зависимости от того, насколько мочевой пузырь наполнен.Какое из следующих утверждений о типах эпителиальных клеток неверно?
- Простые столбчатые эпителиальные клетки выстилают легочную ткань.
- Клетки простого кубического эпителия участвуют в фильтрации крови в почках.
- Псевдослоистые столбчатые эпителии встречаются в одном слое, но расположение ядер создает впечатление, что присутствует более одного слоя.
- Переходный эпителий меняет толщину в зависимости от того, насколько наполнен мочевой пузырь.
Соединительные ткани
Соединительные ткани состоят из матрицы, состоящей из живых клеток и неживого вещества, называемого основным веществом. Основное вещество состоит из органического вещества (обычно белка) и неорганического вещества (обычно минерала или воды). Основной клеткой соединительной ткани является фибробласт. Эта клетка образует волокна, присутствующие почти во всех соединительных тканях. Фибробласты подвижны, способны осуществлять митоз и могут синтезировать любую необходимую соединительную ткань. В некоторых тканях обнаруживаются макрофаги, лимфоциты и иногда лейкоциты. Некоторые ткани имеют специализированные клетки, которых нет в других. матрикс в соединительных тканях придает ткани ее плотность. Когда соединительная ткань имеет высокую концентрацию клеток или волокон, она имеет пропорционально менее плотный матрикс.
Органическая часть или белковые волокна соединительной ткани представляют собой коллагеновые, эластические или ретикулярные волокна. Коллагеновые волокна обеспечивают прочность ткани, предотвращая ее разрыв или отделение от окружающих тканей. Эластические волокна состоят из белка эластина; это волокно может растягиваться на полторы своей длины и возвращаться к своим первоначальным размерам и форме. Эластичные волокна обеспечивают эластичность тканей. Ретикулярные волокна представляют собой третий тип белковых волокон соединительной ткани. Это волокно состоит из тонких нитей коллагена, образующих сеть волокон для поддержки тканей и других органов, с которыми оно связано. Различные типы соединительных тканей, типы клеток и волокон, из которых они состоят, а также расположение образцов тканей приведены в таблице 14.3.
| Ткань | Клетки | Волокна | Местоположение |
|---|---|---|---|
| свободная/ареолярная | фибробласты, макрофаги, некоторые лимфоциты, некоторые нейтрофилы | несколько: коллагеновые, эластические, ретикулярные | вокруг кровеносных сосудов; якоря эпителия |
| плотная волокнистая соединительная ткань | фибробластов, макрофагов, | в основном коллаген | нерегулярный: кожа в норме: сухожилия, связки |
| хрящ | хондроцитов, хондробластов | гиалин: небольшое количество коллагена волокнистый хрящ: большое количество коллагена | скелет акулы, кости плода, человеческие уши, межпозвонковые диски |
| кость | остеобласты, остеоциты, остеокласты | некоторые: коллагеновые, эластичные | скелетов позвоночных |
| жировой | адипоцитов | несколько | жировая ткань (жир) |
| кровь | эритроцитов, | лейкоцитовнет | кровь |
Рыхлая/ареолярная соединительная ткань
Рыхлая соединительная ткань , также называемая ареолярной соединительной тканью, содержит образцы всех компонентов соединительной ткани. Как показано на рис. 14.12, рыхлая соединительная ткань содержит некоторое количество фибробластов; присутствуют также макрофаги. Коллагеновые волокна относительно широкие и окрашиваются в светло-розовый цвет, в то время как эластические волокна тонкие и окрашиваются в темно-синий или черный цвет. Пространство между форменными элементами ткани заполнено матрицей. Содержащийся в соединительной ткани материал придает ей рыхлую консистенцию, похожую на растянутый ватный тампон. Рыхлая соединительная ткань находится вокруг каждого кровеносного сосуда и помогает удерживать сосуд на месте. Ткань также находится вокруг и между большинством органов тела. Таким образом, ареолярная ткань жесткая, но гибкая и состоит из мембран.
Волокнистая соединительная ткань
Волокнистая соединительная ткань содержит большое количество коллагеновых волокон и небольшое количество клеток или матриксного материала. Волокна могут быть расположены неравномерно или регулярно с параллельными нитями. Неравномерно расположенные волокнистые соединительные ткани обнаруживаются в тех областях тела, где нагрузка возникает со всех сторон, например, в дерме кожи. Обычная волокнистая соединительная ткань, показанная на рис. 14.13, содержится в сухожилиях (соединяющих мышцы с костями) и связках (соединяющих кости с костями).
Хрящ
Хрящ представляет собой соединительную ткань с большим количеством матрикса и переменным количеством волокон. Клетки, называемые хондроцитами , образуют матрикс и волокна ткани. Хондроциты находятся в тканевых пространствах, называемых лакунами .
Хрящ с небольшим количеством коллагеновых и эластических волокон представляет собой гиалиновый хрящ, показанный на рис. 14.14. Лакуны беспорядочно разбросаны по всей ткани, а матрикс приобретает молочный или шероховатый вид при обычном гистологическом окрашивании. У акул есть хрящевой скелет, как и почти у всего человеческого скелета на определенной стадии развития до рождения. Остаток этого хряща сохраняется во внешней части человеческого носа. Гиалиновый хрящ также находится на концах длинных костей, уменьшая трение и смягчая сочленения этих костей.
Эластичный хрящ имеет большое количество эластичных волокон, что придает ему невероятную гибкость. Уши большинства позвоночных животных содержат этот хрящ, как и части гортани или голосового аппарата. Фиброхрящ содержит большое количество коллагеновых волокон, придающих ткани огромную прочность. Волокнистый хрящ включает межпозвонковые диски у позвоночных животных. Гиалиновый хрящ, обнаруженный в подвижных суставах, таких как колено и плечо, повреждается в результате старения или травмы. Поврежденный гиалиновый хрящ заменяется волокнистым хрящом, в результате чего суставы становятся «жесткими».
Кость, или костная ткань, представляет собой соединительную ткань, содержащую большое количество матриксного материала двух различных типов. Органический матрикс подобен материалу матрикса, обнаруженному в других соединительных тканях, включая некоторое количество коллагена и эластических волокон. Это придает ткани прочность и гибкость. Неорганическая матрица состоит из минеральных солей, в основном солей кальция, которые придают ткани твердость. Без адекватного органического материала в матрице ткань разрывается; без адекватного неорганического материала в матрице ткань изгибается.
В кости есть три типа клеток: остеобласты, остеоциты и остеокласты. Остеобласты активно участвуют в создании кости для роста и ремоделирования. Остеобласты откладывают костный материал в матрицу, и после того, как матрица окружает их, они продолжают жить, но в сниженном метаболическом состоянии в виде остеоцитов. Остеоциты находятся в костных лакунах. Остеокласты активны в разрушении кости для ее ремоделирования и обеспечивают доступ к кальцию, хранящемуся в тканях. Остеокласты обычно находятся на поверхности ткани.
Кости можно разделить на два типа: компактные и губчатые. Компактная кость находится в теле (или диафизе) длинной кости и на поверхности плоских костей, тогда как губчатая кость находится в конце (или эпифизе) длинной кости. Компактная кость организована в субъединицы, называемые остеонами , как показано на рис. 14.15. Кровеносный сосуд и нерв находятся в центре структуры внутри гаверсова канала, вокруг которого расходящиеся круги лакун, известные как пластинки. Волнистые линии между лакунами — это микроканалы, называемые 9.0003 канальцы ; они соединяют лакуны, чтобы способствовать диффузии между клетками. Губчатая кость состоит из крошечных пластин, называемых трабекулами , эти пластины служат распорками, придающими губчатой кости прочность. Со временем эти пластины могут сломаться, в результате чего кость станет менее упругой. Костная ткань образует внутренний скелет позвоночных животных, обеспечивая структуру животного и точки крепления сухожилий.
Жировая ткань
Жировая ткань или жировая ткань считается соединительной тканью, даже несмотря на то, что она не имеет фибробластов или настоящего матрикса и имеет лишь несколько волокон. Жировая ткань состоит из клеток, называемых адипоцитами, которые собирают и хранят жир в форме триглицеридов для энергетического метаболизма. Жировая ткань дополнительно служит изоляцией, помогая поддерживать температуру тела, позволяя животным быть эндотермическими, и они действуют как защита от повреждений органов тела. Под микроскопом клетки жировой ткани кажутся пустыми из-за выделения жира при обработке материала для просмотра, как видно на рис. 14.16. Тонкие линии на изображении — это клеточные мембраны, а ядра — маленькие черные точки по краям клеток.
Кровь считается соединительной тканью, поскольку она имеет матрицу, как показано на рис. 14.17. Типы живых клеток — это эритроциты (эритроциты), также называемые эритроцитами, и лейкоциты (лейкоциты), также называемые лейкоцитами. Жидкую часть цельной крови, ее матрикс, принято называть плазмой.
Рисунок 14.17. Кровь представляет собой соединительную ткань, имеющую жидкую матрицу, называемую плазмой, и не имеющую волокон. Эритроциты (красные кровяные тельца), преобладающий тип клеток, участвуют в транспорте кислорода и углекислого газа. Также присутствуют различные лейкоциты (лейкоциты), участвующие в иммунном ответе.Клеткой, обнаруженной в крови в наибольшем количестве, являются эритроциты. Эритроциты в образце крови исчисляются миллионами: среднее количество эритроцитов у приматов составляет от 4,7 до 5,5 миллионов клеток на микролитр. Эритроциты постоянно имеют одинаковый размер у вида, но различаются по размеру между видами. Например, средний диаметр эритроцита примата составляет 7,5 мкл, у собаки он близок к 7,0 мкл, а диаметр эритроцитов у кошки составляет 5,9 мкл. Овечьи эритроциты еще меньше – 4,6 мкл. Эритроциты млекопитающих теряют свои ядра и митохондрии, когда они высвобождаются из костного мозга, где они производятся. Красные кровяные тельца рыб, амфибий и птиц сохраняют свои ядра и митохондрии на протяжении всей жизни клетки. Основная функция эритроцита – транспортировать и доставлять кислород к тканям.
Лейкоциты являются преобладающими лейкоцитами в периферической крови. Лейкоциты в крови исчисляются тысячами с измерениями, выраженными в виде диапазонов: количество приматов колеблется от 4800 до 10 800 клеток на мкл, собак от 5 600 до 19 200 клеток на мкл, кошек от 8 000 до 25 000 клеток на мкл, крупного рогатого скота от 4 000 до 12 000 клеток. на мкл, а свиньи от 11 000 до 22 000 клеток на мкл.
Лимфоциты функционируют главным образом в иммунном ответе на чужеродные антигены или материалы. Различные типы лимфоцитов вырабатывают антитела, адаптированные к чужеродным антигенам, и контролируют выработку этих антител. Нейтрофилы являются фагоцитирующими клетками и участвуют в одной из первых линий защиты от микробных захватчиков, способствуя удалению бактерий, проникших в организм. Другим лейкоцитом, обнаруживаемым в периферической крови, является моноцит. Моноциты дают начало фагоцитирующим макрофагам, которые очищают мертвые и поврежденные клетки в организме, независимо от того, являются ли они чужеродными или принадлежат животному-хозяину. Два дополнительных лейкоцита в крови — эозинофилы и базофилы — помогают облегчить воспалительную реакцию.
Слабозернистый материал среди клеток представляет собой цитоплазматический фрагмент клетки костного мозга. Это называется тромбоцитом или тромбоцитом. Тромбоциты участвуют в стадиях, ведущих к свертыванию крови, чтобы остановить кровотечение через поврежденные кровеносные сосуды. Кровь выполняет ряд функций, но в первую очередь она переносит вещества по телу, чтобы доставлять питательные вещества к клеткам и удалять из них отходы.
Мышечные ткани
В телах животных есть три типа мышц: гладкие, скелетные и сердечные. Они различаются наличием или отсутствием исчерченности или полос, количеством и расположением ядер, независимо от того, контролируются ли они вольно или невольно, и их расположением в теле. Таблица 14.4 суммирует эти различия.
| Тип мышц | Полосы | Ядра | Управление | Местоположение |
|---|---|---|---|---|
| гладкая | нет | одинарная, в центре | непроизвольный | внутренние органы |
| скелет | да | много, на периферии | добровольно | скелетные мышцы |
| сердечный | да | одинарная, в центре | непроизвольный | сердце |
Гладкая мышца
Гладкая мышца не имеет исчерченности в своих клетках. Он имеет одно центральное ядро, как показано на рис. 14.18. Сокращение гладкой мускулатуры происходит под непроизвольным контролем вегетативной нервной системы и в ответ на местные условия в тканях. Гладкую мышечную ткань также называют неисчерченной, поскольку она не имеет полосчатого вида скелетных и сердечных мышц. Стенки кровеносных сосудов, трубки пищеварительной системы и трубки половой системы состоят в основном из гладкой мускулатуры.
Скелетная мышца
Скелетная мышца имеет исчерченность клеток, обусловленную расположением сократительных белков актина и миозина. Эти мышечные клетки относительно длинные и имеют несколько ядер по краю клетки. Скелетные мышцы находятся под произвольным контролем соматической нервной системы и находятся в мышцах, которые двигают кости. На рис. 14.18 показана гистология скелетных мышц.
Сердечная мышца
Сердечная мышца, показанная на рис. 14.18, находится только в сердце. Как и скелетная мышца, она имеет поперечную исчерченность в своих клетках, но сердечная мышца имеет одно ядро, расположенное в центре. Сердечная мышца не находится под произвольным контролем, но вегетативная нервная система может влиять на ее ускорение или замедление. Дополнительным признаком клеток сердечной мышцы является линия, которая проходит вдоль конца клетки, когда она упирается в следующую сердечную клетку в ряду. Эта линия называется вставочным диском: она помогает эффективно передавать электрический импульс от одной клетки к другой и поддерживает прочную связь между соседними клетками сердца.
Нервные ткани
Нервные ткани состоят из клеток, специализированных для приема и передачи электрических импульсов от определенных областей тела и для отправки их в определенные места тела. Основной клеткой нервной системы является нейрон, изображенный на рис. 14.19. Крупная структура с центральным ядром является телом нейрона. Отростки тела клетки представляют собой либо дендриты, специализирующиеся на получении входных данных, либо отдельные аксоны, специализирующиеся на передаче импульсов. Также показаны некоторые глиальные клетки. Астроциты регулируют химическую среду нервной клетки, а олигодендроциты изолируют аксон, благодаря чему электрический нервный импульс передается более эффективно. Другие глиальные клетки, которые не показаны, поддерживают потребности нейрона в питании и отходах. Некоторые из глиальных клеток являются фагоцитами и удаляют остатки или поврежденные клетки из ткани. Нерв состоит из нейронов и глиальных клеток.
Концепция в действии
Нажмите на интерактивный обзор, чтобы узнать больше об эпителиальных тканях.
Патологоанатом
Патологоанатом — это врач или ветеринар, специализирующийся на лабораторном выявлении заболеваний у животных, в том числе у людей. Эти специалисты заканчивают медицинское образование, а затем проходят последипломную ординатуру в медицинском центре. Патолог может наблюдать за клиническими лабораториями для оценки тканей тела и образцов крови для выявления заболеваний или инфекций. Они исследуют образцы тканей под микроскопом для выявления рака и других заболеваний. Некоторые патологоанатомы проводят вскрытие, чтобы определить причину смерти и прогрессирование заболевания.
Глоссарий
хрящ: тип соединительной ткани с большим количеством матрикса основного вещества, клетками, называемыми хондроцитами, и некоторым количеством волокон
хондроцит: клетка, обнаруженная в хряще выше, чем в ширину, специализируются на абсорбции
соединительная ткань: тип ткани, состоящей из клеток, матрикса основного вещества и волокон
кубовидный эпителий: эпителий, состоящий из клеток кубической формы, специализирующийся на железистых функциях
полость спины: полость тела на задней или задней части животного; включает полость черепа и позвоночника
эктотерм: животное, неспособное поддерживать относительно постоянную внутреннюю температуру тела
эндотерм: животное, способное поддерживать относительно постоянную внутреннюю температуру тела
эпителиальная ткань: ткань, которая либо выстилает, либо покрывает органы или другие ткани
активация: оцепенение в ответ на экстремально высокие температуры и недостаток воды
волокнистая соединительная ткань: тип соединительной ткани с высокой концентрацией волокон
гибернация: оцепенение в течение длительного периода времени, например зимой
гомеостаз: динамическое равновесие, поддержание соответствующих функций организма
лакуна: пространство в хрящах и костях, содержащее живые клетки
матрикс: компонент соединительной ткани, состоящий как из живых, так и из неживых (основных веществ) клеток
петля отрицательной обратной связи: обратная связь с механизмом управления, который увеличивает или уменьшает стимул вместо его сохранения
остеон: субъединица компактной кости
петля положительной обратной связи: обратная связь с механизмом управления, который продолжает направление стимула
псевдостратифицированный: слой эпителия, который кажется многослойным, но представляет собой простое покрытие
заданная точка: средняя или целевая точка гомеостаза
простой эпителий: один слой эпителиальных клеток
плоскоклеточный эпителий: тип эпителия, состоящий из плоских клеток, специализирующийся на содействии диффузии или предотвращении истирания
многослойный эпителий: несколько слоев эпителиальных клеток
оцепенение: снижение активности и метаболизма, которое позволяет животному выживать в неблагоприятных условиях
трабекула: крошечная пластинка, образующая губчатую кость и придающая ей прочность
эпителиальных клеток | Спросите у биолога
показать/скрыть слова, чтобы узнать
Гормоны: химические вещества, выделяемые клетками или железами тела.
Молекула: химическая структура, состоящая из двух или более атомов, связанных друг с другом химической связью. Вода представляет собой молекулу, состоящую из двух атомов водорода и одного атома кислорода (h3O)… подробнее
Рецептор: молекула на поверхности клетки, которая реагирует на определенные молекулы и получает химические сигналы, посылаемые другими клетками.
Секрет: для высвобождения любого вещества, молекулы или химического вещества из железы или клетки тела.
вернуться к комиксу
Эпителиальные клетки
Где эпителиальные клетки?
Взгляните на кожу рук. Даже если вы думаете, что ваша кожа представляет собой единую гладкую поверхность, на самом деле она состоит из миллионов эпителиальных клеток, которые плотно прилегают друг к другу.
Это не единственное место, где вы найдете эти клетки. Эпителиальные клетки также выстилают внутреннюю часть вашего горла, кишечника, кровеносных сосудов и всех ваших органов. Они являются барьером между внутренней и внешней частью вашего тела и часто являются первым местом, которое атакуют вирусы, поскольку они начинают свое проникновение глубже в тело.
Что делают эпителиальные клетки?
Эпителиальные клетки являются защитным экраном организма. Взгляните еще раз на свою руку. Он покрыт эпителиальными клетками, которые защищают ваше тело, являясь барьером между вашими внутренними клетками и грязью и микробами в окружающей среде. Они также могут растягиваться, поэтому вы можете двигать пальцами и руками во многих положениях. Вы также можете поблагодарить свои эпителиальные клетки за выделение пота, который охлаждает вас, когда вы тренируетесь или когда на улице жарко. Чтобы узнать больше о своей коже и о том, как она работает каждый день, послушайте этот подкаст.
Окрашенные эпителиальные клетки по Page Baluch.
Другие эпителиальные клетки помогают вам воспринимать окружающую среду благодаря специальным сенсорам, называемым рецепторами, которые собирают сигналы. Когда вы пробуете любимую еду или чувствуете запах цветка, рецепторы в этих клетках посылают сигнал в ваш мозг, чтобы вы могли наслаждаться каждым кусочком и сладким запахом.
Когда вы проглатываете кусок пищи, он движется по пути, выстланному эпителиальными клетками. Когда он попадает в ваш кишечник, другой набор эпителиальных клеток поглощает и переносит питательные вещества из продуктов, которые вы едите, и помогает перерабатывать их для получения энергии, которую может использовать ваше тело. Преобразование энергии пищи в энергию, которую может использовать ваше тело, — это работа молекул, называемых ферментами. Опять же, это эпителиальные клетки, которые производят и выделяют ферменты в вашем желудке. Эпителиальные клетки также выделяют гормоны в ваши кровеносные сосуды, слизь в носу и грудное молоко, которым матери кормят своих детенышей.
Как выглядят эпителиальные клетки?
Если вы внимательно посмотрите на эпителиальные клетки под микроскопом, вы увидите, что они плотно упакованы вместе. Это помогает создать защитный барьер для нашего тела. Между каждой эпителиальной клеткой есть также специальные дверные соединения, называемые щелевыми соединениями. Щелевые контакты — это место, где клетки обмениваются питательными веществами. К сожалению, иногда вирусы могут использовать эти двери для распространения и между клетками!
Эпителиальные клетки бывают разной формы в зависимости от того, в какой части тела они находятся. Эти формы называются плоскоклеточными, кубовидными, столбчатыми и реснитчатыми столбчатыми.
Клетки плоского эпителия являются плоскими и обычно выстилают поверхности, требующие беспрепятственного тока жидкости, например, кровеносные сосуды. Они также выстилают области, которые требуют очень тонкой поверхности для прохождения молекул, например, воздушные мешочки в ваших легких. | |
| Кубовидные эпителиальные клетки , как следует из их названия, имеют форму кубов. Обычно они обнаруживаются в тканях, которые выделяют или поглощают вещества, например, в почках и железах. | |
| Столбчатые эпителиальные клетки длинные и тонкие, как столбики. Они обычно находятся в местах, выделяющих слизь, таких как желудок. Они также могут специализироваться на получении сенсорной информации в таких местах, как вкусовые рецепторы на вашем языке и внутри вашего носа. | |
| Реснитчатые столбчатые клетки имеют свою апикальную (или обращенную наружу) поверхность, покрытую множеством крошечных волосков, называемых ресничками. Они используются для проталкивания слизи и других частиц, заставляя их течь в определенном направлении. |
В дополнение к этим формам эпителиальные клетки могут быть описаны как простые или многослойные. Эти термины относятся к тому, сколько слоев присутствует. Простая ткань имеет только один слой эпителиальных клеток, тогда как многослойная ткань имеет много слоев, наложенных друг на друга. Многослойные клетки находятся в местах, которые должны выдерживать большой износ от окружающей среды.
Примером может служить ваша кожа, состоящая из множества многослойных слоев эпителиальных клеток. По мере того, как верхний слой изнашивается, клетки из нижних слоев постоянно вырастают, чтобы заменить его.
вернуться к комиксу
Подробнее о: Вирусная атака
Взаимосвязь между свойствами поверхностного, пограничного слоя и столбчатого аэрозоля, полученная летом и ранней осенью над континентальной городской территорией в Варшаве, Польша
Альтаратц, О., Бар-Ор, Р. З., Воллнер, У. ., и Корень, И.: Относительная влажность и его влияние на оптическую толщину аэрозоля в окрестности конвективных облаков, Environ. Рез. Лет., 8, 034025, г. https://doi.org/10.1088/1748-9326/8/3/034025, 2013.
Аладос-Арболедас, Л., Мюллер, Д., Герреро-Раскадо, Дж.,
Навас-Гусман Ф., Перес-Рамирес Д. и Олмо Ф.: Оптика
и микрофизические свойства свежего аэрозоля горения биомассы, извлеченного
рамановский лидар, звездная и солнечная фотометрия,
Геофиз. Рез. Lett., 38, L01807, https://doi.org/10.1029/2010GL045999, 2011.
Амиридис В., Балис Д., Казадзис С., Баис А., Гианнакаки Э., Папаяннис, А. и Зерефос, К.: Четырехлетний аэрозоль наблюдения с помощью рамановского лидара в Салониках, Греция, в рамках Европейская сеть лидарных исследований аэрозолей (EARLINET), J. Geophys. Рез., 110, D21203, https://doi.org/10.1029/2005JD006190, 2005.
Амиридис, В., Балис, Д.С., Гианнакаки, Э., Штоль, А., Казадзис, С., Кукули, М.Е., и Занис, П.: Оптические характеристики аэрозолей горения биомассы над Юго-Восточной Европой, определенные по УФ-излучению. — Лидарные измерения комбинационного рассеяния света, Атмос. хим. Phys., 9, 2431–2440, https://doi.org/10.5194/acp-9-2431-2009, 2009. о пыли в воздухе, Геогр. Анн., 12, 156–166, https://doi.org/10.1080/20014422.1929.11880498, 1929.
Ансманн, А., Теше, М., Книпперц, П., Бирвирт, Э., Альтхаузен, Д.,
Мюллер, Д., и Шульц, О.: Вертикальное профилирование
конвективные пылевые шлейфы в Южном Марокко во время SAMUM, Tellus B, 61, 340–353, https://doi. org/10.1111/j.1600-0889.2008.00384.x, 2009.
Ансманн, А., Баарс, Х. ., Чудновский А., Маттис И., Веселовский И., Хаариг М., Зайферт П., Энгельманн Р. и Вандингер У.: Экстремальные уровни дыма канадских лесных пожаров в стратосфере над Центральной Европой 21–22 августа 2017 г., Атмос. хим. Phys., 18, 11831–11845, https://doi.org/10.5194/acp-18-11831-2018, 2018.
Баарс, Х., Каниц, Т., Энгельманн, Р., Альтхаузен, Д., Хиз, Б., Комппула, М., Прейслер, Дж., Теше , М., Ансманн, А., Вандингер, У., Лим, Дж.-Х., Ан, Дж.Ю., Стахлевска, И.С., Амиридис, В., Марину, Э., Зайферт, П., Хофер, Дж., Скупин А., Шнайдер Ф., Больманн С., Фот А., Блей С., Пфюллер А., Гианнакаки Э., Лихавайнен Х., Вийсанен Ю., Худа Р. К., Перейра , С. Н., Бортоли Д., Вагнер Ф., Маттис И., Яницка Л., Маркович К. М., Ахтерт П., Артаксо П., Пауликвис Т., Соуза Р. А. Ф., Шарма В. П., ван Зил, П. Г., Бьюкес, Дж. П., Сан, Дж., Ровер, Э. Г., Денг, Р., Мамури, Р.-Э., и Саморано, Ф.: Обзор первого десятилетия PollyNET: развивающаяся сеть автоматизированные лидары рамановской поляризации для непрерывного профилирования аэрозолей, Atmos. хим. Phys., 16, 5111–5137, https://doi.org/10.5194/acp-16-5111-2016, 2016.
Баарс, Х., Ансманн, А., Онайзер, К., Хаариг, М., Энгельманн, Р., Альтхаузен, Д., Ханссен, И., Гауса М., Петручук А., Шкоп А., Стахлевска И.С., Ван Д., Райххардт Дж., Скупин А., Маттис И., Трикл Т., Фогельманн Х., Навас- Гусман Ф., Хефеле А., Ачесон К., Рут А. А., Татаров Б., Мюллер Д., Ху К., Подвин Т., Голуб П., Веселовский И., Пьетрас , C., Haeffelin, M., Fréville, P., Sicard, M., Comeron, A., Fernández García, A.J., Molero Menéndez, F., Córdoba-Jabonero, C., Guerrero-Rascado, J.L., Alados- Арболедас Л., Бортоли Д., Коста М.Дж., Диониси Д., Либерти Г.Л., Ван Х., Саннино А., Папагианнопулос Н., Боселли А., Мона Л., Д’ Амико Г., Романо С., Перроне М. Р., Белеганте Л., Николае Д., Григоров И., Гиалитаки А., Амиридис В., Супиона О., Папаяннис А., Мамури , Р.-Э., Нисанци А., Хиз Б., Хофер Дж., Шехнер Ю.Ю., Вандингер У. и Паппалардо Г.: Беспрецедентный стратосферный дым 2017–2018 гг. событие: Фаза распада и свойства аэрозоля, наблюдаемые с помощью EARLINET, Atmos. хим. физ. Обсудить., https://doi.org/10.5194/acp-2019-615, обзор, 2019 г.
Барлаж М., Мяо С. и Чен Ф.: Влияние физических параметризации на прогноз погоды с высоким разрешением в течение двух Китайские мегаполисы // J. Geophys. рез., 121, 4487–4498, https://doi.org/10.1002/2015JD024450, 2016.
Беннуна, Ю., Качорро, В.Е., Матеос, Д., Бургос, М.А., Толедано, К., Торрес, Б., и де Фрутос, А.: Долгосрочные сравнительное исследование столбчатой и поверхностной массовой концентрации аэрозоля свойств в фоновом режиме, Atmos. Окружающая среда., 140, 261–272, https://doi.org/10.1016/j.atmosenv.2016.05.061, 2016.
Бегейн С., Аллери С., Вацлавчик С. и Позорски Ю.: Применение Динамические системы на основе POD для рассеивания и осаждение частиц в турбулентном канальном течении, Int. J. Многофазный поток, 58, 97–113, https://doi.org/10.1016/j.ijmultiphaseflow.2013.09.001, 2014.
Бергин, М. Х., Шварц, С. Э., Халтор, Р. Н., Огрен, Дж. А., и Главка,
Д. Л.: Сравнение рассчитанной оптической толщины аэрозоля. приземные измерения с данными, определенными фотометрией Солнца для безоблачного
условия на континентальной территории США, J. Geophys.
Рез., 105, 6807–6816, https://doi.org/10.1029/1999JD
Биниетоглу, И., Басарт, С., Аладос-Арболедас, Л., Амиридис, В., Аргирули, А., Баарс, Х., Балдасано, Дж. М., Балис, Д., Белеганте, Л., Браво-Аранда, Дж. А., Бурлицци, П., Карраско, В., Чайковский, А., Комерон, А., Д’Амико, Г., Филиоглу, М., Гранадос-Муньос, М. Х., Герреро-Раскадо , Дж. Л., Илич, Л., Коккалис, П., Маурици, А., Мона, Л., Монти, Ф., Муньос-Поркар, К., Николае, Д., Папаяннис, А., Паппалардо, Г., Пеянович Г., Перейра С. Н., Перроне М. Р., Петручук А., Посиняк М., Рокаденбош Ф., Родригес-Гомес А., Сикард М., Сиомос Н., Шкоп А., Терраделлас, Э., Цекери, А., Вукович, А., Вандингер, У. и Вагнер, Дж.: Методология исследования производительности модели пыли с использованием синергетического извлечения концентрации пыли EARLINET/AERONET, Atmos. Изм. Тех., 8, 3577–3600, https://doi. org/10.5194/amt-8-3577-2015, 2015.
Бёкманн, К., Вандингер, У., Ансманн, А., Босенберг, Дж., Амиридис, В., Боселли А., Делаваль А., Де Томази Ф., Фриуд, М., Григоров И.В.: Взаимное сравнение аэрозольных лидаров в рамках проект ЭРЛИНЕТ. 2. Обратное аэрозольное рассеяние алгоритмы, прил. Опт., 43, 977–989, https://doi.org/10.1364/AO.43.000977, 2004.
Бёкманн, К., Миронова, И., Мюллер, Д., Шнайденбах, Л., и Несслер, Р.: Микрофизические параметры аэрозоля из многоволнового диапазона. лидар, J. Opt. соц. Являюсь. А, 22, 518–528, https://doi.org/10.1364/JOSAA.22.000518, 2005 г.
Бонн, Б., фон Шнайдемессер, Э., Андрих, Д., Кведенау, Дж., Гервиг, Х., Людеке, А., Кура, Й., Питч, А., Элерс, К., Клемп, Д., Кофаль К., Нотхард Р., Кершбаумер А., Юнкерманн В., Гроте Р., Поль Т., Вебер К., Лоде Б., Шенбергер П., Чуркина Г. Г., Батлер, Т. М., и Лоуренс, М. Г.: BAERLIN 2014 – влияние типов земной поверхности на горизонтальную неоднородность уровней загрязнителей воздуха в Берлине, Atmos. хим. Phys., 16, 7785–7811, https://doi.org/10.5194/acp-16-7785-2016, 2016.
Бертон, С. П., Ферраре, Р. А., Хостетлер, К. А., Хейр, Дж. В., Роджерс, Р. Р., Обланд, М. Д., Батлер, К. Ф., Кук, А. Л., Харпер, Д. Б., и Фройд, К. Д.: Классификация аэрозолей с использованием бортового высокого спектрального разрешения Лидарные измерения – методология и примеры, Atmos. Изм. Tech., 5, 73–98, https://doi.org/10.5194/amt-5-73-2012, 2012.
Burton, S.P., Hair, J.W., Kahnert, M., Ferrare, R.A., Hostetler, К. А., Кук А. Л., Харпер Д. Б., Беркофф Т. А., Симэн С. Т., Коллинз Дж. Э., Фенн М. А. и Роджерс Р. Р.: Наблюдения за спектральной зависимостью коэффициента линейной деполяризации частиц аэрозолей с использованием высокого спектрального разрешения НАСА в Лэнгли. Лидар, Атмос. хим. Phys., 15, 13453–13473, https://doi.org/10.5194/acp-15-13453-2015, 2015.
Чен, Б. и Кан, Х.: Загрязнение воздуха и здоровье населения: глобальный
вызов, Окружающая среда. Здоровье Пред. мед., 13, 94–101,
https://doi. org/10.1007/s12199-007-0018-5, 2008.
Ченг, Ю. Ф., Виденсолер, А., Эйхлер, Х., Хайнценберг, Дж., Теше, М., Ансманн, А., Вендиш, М., Су, Х., Альтхаузен, Д., Херрманн Х., Гнаук Т., Брюггеманн Э., Ху М. и Чжан Ю. Х.: Зависимость аэрозоля от относительной влажности оптические свойства и прямое радиационное воздействие на границе поверхности слой в Синькене в дельте Жемчужной реки в Китае: численное исследование на основе наблюдений, Atmos. Окружающая среда, 42, 6373–6397, https://doi.org/10.1016/j.atmosenv.2008.04.009, 2008.
Чилинский М.Т., Маркович К.М., Завадская О., Стахлевская И.С., Кумала В., Петельски Т., Макух П., Вестфаль Д. Л. и Загаевски Б.: Моделирование и наблюдение минеральной пыли в оптическом диапазоне Недвижимость в Центральной Европе, Acta Geophys., 64, 2550–2590, https://doi.org/10.1515/acgeo-2016-0069, 2016.
Комерон, А., Сикард, М., и Рокаденбош, Ф.: Корреляция вейвлетов
Метод преобразования и метод градиента для определения
Наслоение аэрозолей по результатам лидара: некоторые комментарии, J. Atmos. Океан. Тех.,
30, 1189–1193, https://doi.org/10.1175/JTECH-D-12-00233.1, 2013 г.
Коста-Сурос М., Стахлевска И.С., Немук А., Талиану С., Хиз Б., и Энгельманн, Р.: Исследование воздушной массы модификация над Польшей и Румынией, наблюдаемая с помощью многоволновые лидары рамановской деполяризации, 27-е место Международная конференция по лазерным радарам, Нью-Йорк, США, 5–10 июля 2015 г., стр. 1–4, 2015.
Данг Р., Ян Ю., Ху Х.-М., Ван З. и Чжан С.: Обзор методов для диагностики пограничного слоя атмосферы Высота (ABLH) с использованием данных аэрозольного лидара, дистанционное зондирование, 11, 1590, https://doi.org/10.3390/rs11131590, 2019.
Доусон, К.В., Месхидзе, Н., Джоссет, Д., и Гассо, С.: Космические наблюдения лидарного отношения морских аэрозолей, Atmos. хим. Phys., 15, 3241–3255, https://doi.org/10.5194/acp-15-3241-2015, 2015.
Деланоэ, Дж. и Хоган, Р. Дж.: Вариант схемы извлечения льда
свойства облаков от комбинированного радара, лидара и инфракрасного излучения
радиометр, J. Geophys. Рез., 113, D07204, https://doi.org/10.1029/2007JD009000, 2008.
Де Леу, Ф., Слейтер, Р., ван Брейгель, П., и Богман, Ф.: Загрязнение воздуха озона в Европе в 1999 и лето 2000 г., Тематический отчет Европейского агентства по охране окружающей среды № 1/2001, ЕАОС, Копенгаген, Дания, 2001.
Ди Бьяджо, К., Пелон, Дж., Анселлет, Г., Базуро, А., и Мариаж, В.: Источники, нагрузка, вертикальное распределение и судьба зимнего времени аэрозоль к северу от Шпицбергена по объединенным данным V4 CALIOP, наземный IAOOS лидарные наблюдения и анализ траекторий, Дж. Геофиз. Рез.-Атмос., 123, 1363–1383, https://doi.org/10.1002/2017JD027530, 2018.
Дорнбрак А., Стахлевска И.С., Риттер К. и Нойбер Р.: Распределение аэрозолей вокруг Шпицбергена во время сильных восточных ветров, Atmos. хим. Phys., 10, 1473–149.0, https://doi.org/10.5194/acp-10-1473-2010, 2010.
Du, C., Liu, S., Yu, X., Li, X., Chen, C., Peng , Ю., Донг, Ю., Донг, З. и
Ван, Ф.: Высота городского пограничного слоя
характеристики и взаимосвязь с массовой концентрацией твердых частиц
в Сиане, Центральный Китай, аэрозоль Air Qual. Res., 13, 1598–1607, https://https://doi.org/10.4209/aaqr.2012.10.0274, 2013.
Engelmann, R., Kanitz, T., Baars, H., Heese, Б., Альтхаузен Д., Скупин А., Вандингер У., Комппула М., Стахлевска И.С., Амиридис В., Марину Э., Маттис И., Линне Х. и Ансманн, A.: Автоматизированная многоволновая комбинационная поляризация и лидар водяного пара PollyXT: новое поколение, Atmos. Изм. Тех., 9, 1767–1784, https://doi.org/10.5194/amt-9-1767-2016, 2016.
Фан, Дж., Ван, Ю., Розенфельд, Д., и Лю, X.: Обзор аэрозольное облако взаимодействия: механизмы, значение и проблемы, Дж. Атмос. Sci., 73, 4221–4252, https://doi.org/10.1175/JAS-D-16-0037.1, 2016.
Feingold, G., McComiskey, A., Yamaguchi, T., Johnson, J. , Карслоу, К., и Шмидт, К.С.: Новые подходы к количественному определению аэрозолей влияние на радиационное действие облаков, П. Нац. акад. науч. США, 113, 5812–5819, https://doi.org/10.1073/pnas.1514035112, 2016.
Fiebig, M., Stohl, A., Wendisch, M., Eckhardt, S. , and Petzold, A.: Зависимость солнечного радиационного воздействия аэрозоля лесных пожаров на старение и состояние смеси, атм. хим. Phys., 3, 881–891, https://doi.org/10.5194/acp-3-881-2003, 2003.
Филип, Л. и Стефан, С.: Изучение корреляции между приземными Массовая концентрация PM10 и оптическая аэрозольность Глубина, Дж. Атмос. Соль.-Терр. Phys., 73, 1883–1889, https://doi.org/10.1016/j.jastp.2011.04.027, 2011.
Флентье, Х., Хиз, Б., Рейхардт, Дж. и Томас, В.: Профилирование аэрозолей с использованием сети облакомеров Немецкой метеорологической службы, Atmos. Изм. Тех. Discuss., 3, 3643–3673, https://doi.org/10.5194/amtd-3-3643-2010, 2010.
Freudenthaler, V., Esselborn, M., Wiegner, M., Heese, B. , Теще, М.,
Ансманн А., Мюллер Д., Альтхаузен Д., Вирт М.,
Фикс А., Эрет Г., Книпперц П., Толедано К., Гастайгер Дж., Гархаммер,
М. и Зеефельднер М.: Коэффициент деполяризации
профилирование на нескольких длинах волн в чистой сахарской пыли во время SAMUM 2006,
Теллус Б, 61, 165–179,
https://https://doi. org/10.1111/j.1600-0889.2008.00396.x, 2009.
Фройденталер В., Линне Х., Чайковский А., Рабус Д. и Грос , S.: Инструменты обеспечения качества лидара EARLINET, Atmos. Изм. Тех. Обсудить., https://doi.org/10.5194/amt-2017-395, в обзоре, 2018 г.
Фот, А., Каниц, Т., Энгельманн, Р., Баарс, Х., Раденц, М. , Seifert, P., Barja, B., Fromm, M., Kalesse, H. и Ansmann, A.: Вертикальное распределение аэрозолей в средних широтах южного полушария, наблюдаемое с помощью лидара в Пунта-Аренас, Чили (53,2 ∘ ю.ш. и 70,9 ∘ з.д.), во время АЛЬПАКА, Атмос. хим. Phys., 19, 6217–6233, https://doi.org/10.5194/acp-19-6217-2019, 2019.
Fuzzi, S., Baltensperger, U., Carslaw, K., Decesari, S. , Деньер ван дер Гон, Х., Факкини, М.С., Фаулер, Д., Корен, И., Лэнгфорд, Б., Ломанн, У., Немиц, Э., Пандис, С., Рийпинен, И., Рудич, Ю., Шаап М., Словик Дж. Г., Спраклен Д. В., Виньяти Э., Уайлд М., Уильямс М. и Джилардони С.: Твердые частицы, качество воздуха и климат: извлеченные уроки и будущие потребности , Атмос. хим. Phys., 15, 8217–8299, https://doi.org/10.5194/acp-15-8217-2015, 2015.
Гастайгер, Дж. и Фройденталер, В.: Преимущество коэффициента деполяризации при λ = 1064 нм для получения микрофизики аэрозоля по лидарным измерениям, атм. Изм. Tech., 7, 3773–3781, https://doi.org/10.5194/amt-7-3773-2014, 2014.
Гаятри, К., Патаде, С., и Прабха, Т.В.: Взаимодействие аэрозоля и облака в глубокие конвективные облака над Индийским полуостровом с помощью спектральная (бинарная) микрофизика, J. Atmos. наук, 74, 3145–3166, https://doi.org/10.1175/JAS-D-17-0034.1, 2017 г.
Гайсс, А., Вигнер, М., Бонн, Б., Шефер, К., Форкель, Р., фон Шнайдемессер, Э., Мюнкель, К., Чан, К. Л., и Нотхард, Р.: Смесительный слой высота как показатель качества городского воздуха?, Атмос. Изм. Tech., 10, 2969–2988, https://doi.org/10.5194/amt-10-2969-2017, 2017.
Гианнакаки Э., Балис Д. С., Амиридис В. и Зерефос К. : Оптические свойства различных типов аэрозолей: семь лет комбинированных лидарных измерений комбинационного упругого обратного рассеяния в Салониках, Греция, Atmos. Изм. Тех., д. 3, 569–578, https://doi.org/10.5194/amt-3-569-2010, 2010.
Ган, С.Дж., Ван, М., Чжан, С., Феррачат, С., Геттельман, А., Грисфеллер , Дж., Киплинг З., Ломанн У., Моррисон Х., Нойбауэр, Д., Партридж Д.Г., Стиер П., Такемура Т., Ван Х. и Чжан К.: Проблемы ограничения воздействия антропогенных аэрозолей о радиационном воздействии облаков с использованием современной пространственно-временной изменчивости, П. Натл. акад. науч. USA, 113, 5804–5811, https://doi.org/10.1073/pnas.1514036113, 2016.
Джайлз Д. М., Синюк А., Сорокин М. Г., Шафер Дж. С., Смирнов А., Слуцкер, И., Экк, Т.Ф., Холбен, Б.Н., Льюис, Дж.Р., Кэмпбелл, Дж.Р., Велтон, Э.Дж., Коркин, С.В., и Ляпустин, А.И.: Достижения в базе данных Aerosol Robotic Network (AERONET) Версия 3 – автоматизированная, почти реальная алгоритм контроля качества времени с улучшенным скринингом облаков для измерений оптической толщины аэрозоля (AOD) солнечного фотометра, Atmos. Изм. Тех., 12, 169–209, https://doi.org/10. 5194/amt-12-169-2019, 2019.
Гранадос-Муньос, М. Дж., Навас-Гусман, Ф., Браво-Аранда, Дж. А., Герреро-Раскадо, Дж. Л., Лямани, Х., Валенсуэла, А., Титос, Г., Фернандес-Гальвес, Х., и Аладос-Арболедас, Л.: Гигроскопический рост частиц атмосферного аэрозоля на основе активных измерений дистанционного зондирования и радиозондирования: отдельные случаи на юго-востоке Испании , Атмос. Изм. Tech., 8, 705–718, https://doi.org/10.5194/amt-8-705-2015, 2015.
Groß, S., Tesche, M., Freudenthaler, V., Toledano, C. , Вигнер, М., Ансманн А., Альтхаузен Д. и Зеефельднер М.: Характеристика сахарской пыли, морского аэрозоля и смесей аэрозоль и пыль, сжигающие биомассу, с помощью многоволновой деполяризации и измерений рамановского лидара во время SAMUM 2, Теллус Б, 63 года, 706–724, https://doi.org/10.1111/j.1600-0889..2011.00556.x, 2011.
Гросс, С., Эссельборн, М., Вайнцирль, Б., Вирт, М., Фикс, А. и Петцольд, А.: Классификация аэрозолей с помощью бортовых лидарных наблюдений с высоким спектральным разрешением, Атмос. хим. Phys., 13, 2487–2505, https://doi.org/10.5194/acp-13-2487-2013, 2013.
Groß, S., Freudenthaler, V., Schepanski, K., Toledano, C. , Шефлер, А., Ансманн, А., и Вайнцирль, Б.: Оптические свойства переносимой на большие расстояния сахарской пыли над Барбадосом, измеренные с помощью двухволновых лидарных измерений комбинационного рассеяния деполяризации, Atmos. хим. Phys., 15, 11067–11080, https://doi.org/10.5194/acp-15-11067-2015, 2015.
Грунд, CJ и Элоранта, EW: Университет Висконсина High Spectral Разрешение Лидар, Опт. Eng., 30, 6–12, 1991.
Го, Х., Ван, Ю., и Чжан, Х.: Характеристика критериев воздуха загрязняющих веществ в Пекине в 2014–2015 гг., Environ. рез., 154, 334–344, https://doi.org/10.1016/j.envres.2017.01.029, 2017.
Го, Ж.-П., Чжан, X.-Y., Че, Х.-З. ., Гун С.-Л., Ан X., Цао С.-X.,
Гуанг Дж., Чжан Х., Ван Ю.-К., Чжан Х.-К.,
Сюэ, М., и Ли, X.-W.: Корреляция между концентрациями ТЧ и аэрозолем
оптическая толщина в восточном Китае, атм.
Окружающая среда, 43, 5876–5886, https://doi. org/10.1016/j.atmosenv.2009.08.026,
2009.
Хаариг, М., Ансманн, А., Гастайгер, Дж., Кандлер, К., Альтхаузен, Д., Баарс, Х., Раденц М. и Фаррелл Д. А.: Сухой против Оптические свойства влажных морских частиц: RH-зависимость деполяризации отношение, обратное рассеяние и поглощение многоволновые лидарные измерения во время SALTRACE, Atmos. хим. физ., 17, 14199–14217, https://doi.org/10.5194/acp-17-14199-2017, 2017.
Хаариг М., Ансманн А., Баарс Х., Хименес С., Веселовский И. , Энгельманн Р. и Альтхаузен Д.: Коэффициенты деполяризации и лидара на длинах волн 355, 532 и 1064 нм и микрофизические свойства старого тропосферного и стратосферного дыма лесных пожаров в Канаде, Atmos. хим. Phys., 18, 11847–11861, https://doi.org/10.5194/acp-18-11847-2018, 2018.
Хеффелин М., Анджелини Ф., Морилль Ю., Мартуччи Г., Фрей С., Гобби Г.
П., Лолли С., О’Дауд С.Д., Соваж Л., Сюереф Реми,
И., Вастин Б. и Файст Д. Г.: Оценка извлечения высоты смешивания
от автоматических профилирующих лидаров и облакомеров в поле зрения
будущих интегрированных сетей в Европе, Bound. -Lay. Метеорол., 143, 49–75,
https://doi.org/10.1007/s10546-011-9643-z, 2012.
Харрисон, Л., Михальский, Дж., и Берндт, Дж.: Автоматическое вращение мультифильтра радиометр теневого диапазона: прибор для измерения оптической глубины и радиации, Прикл. Опт., 33, 5118–5125, г. https://doi.org/10.1364/AO.33.005118, 1994.
He, Q., Li, C., Mao, J., Lau, A.K.H., and Chu, D.: Анализ аэрозолей вертикальное распределение и изменчивость в Гонконге, J. Геофиз. Res., 113, D14211, https://doi.org/10.1029/2008JD009778, 2008.
Heese, B. and Wiegner, M.: Вертикальные аэрозольные профили, полученные методом комбинационного рассеяния. поляризационные лидарные наблюдения в сухой сезон Полевая кампания AMMA, J. Geophys. Рез., 113, D00C11, https://doi.org/10.1029/2007JD009487, 2008.
Холбен, Б. Н., Эк, Т. Ф., Слуцкер, И., Танре, Д., Буис, Дж. П., Сетцер, А.,
Вермоте Э., Рейган Дж. А., Кауфман Ю. Дж.,
Накжима Т., Лавеню Ф., Янковяк И. и Смирнов А.: AERONET – A
объединенная сеть приборов и архив данных для
характеристика аэрозолей, Remote Sens. Environ., 66, 1–16,
https://doi.org/10.1016/S0034-4257(98)00031-5, 1998.
Хорват, Х., Аладос Арболедас, Л., и Олмо Рейес, Ф.Дж.: Угловое рассеяние пылевого аэрозоля Сахары, Atmos. хим. Phys., 18, 17735–17744, https://doi.org/10.5194/acp-18-17735-2018, 2018.
Ху К., Голуб П., Веселовский И., Браво-Аранда, Дж.-А., Поповичи И.Е., Подвин Т., Хаффелин М., Лопатин А., Дубовик О., Пьетрас С., Хуанг X., Торрес Б. и Чен С. : Канадские дымовые шлейфы, переносимые на большие расстояния, в нижних слоях стратосферы над северной Францией, Атмос. хим. физ., 19, 1173–1193, https://doi.org/10.5194/acp-19-1173-2019, 2019.
Хатчисон, К. Д., Фаруки, С. Дж., и Смит, С.: Улучшение корреляций между оптической толщиной аэрозоля MODIS и наземные PM 2,5 наблюдений с помощью трехмерного пространственного анализа, Атмос. Environ., 42, 530–543, https://doi.org/10.1016/j.atmosenv.2007.09.050, 2008.
Ярлори, М., Мадонна, Ф., Ризи, В., Трикл, Т. и Амодео, А.: Концепции эффективного разрешения для лидарных наблюдений, Atmos. Изм. Тех., 8, 5157–5176, https://doi.org/10.5194/amt-8-5157-2015, 2015.
Иллингворт, А. Дж., Баркер, Х. В., Белджаарс, А., Чекальди, М., Чепфер, Х., Клербо, Н., Коул, Дж., Деланоэ, Дж., Доменек, К., Донован Д. П., Фукуда С., Хираката М., Хоган Р. Дж., Хюнербейн А., Коллиас П., Кубота Т., Накадзима Т., Накадзима Т. Ю., Нисидзава Т., Оно Ю., Окамото Х., Оки Р., Сато К., Сато М., Шепард М., Веласкес-Бласкес А., Вандингер У., Вер, Т., и ван Задельхофф, Г.-Дж.: Спутник EarthCARE: следующий шаг вперед в глобальных измерениях облаков, аэрозолей, осадки и радиация, Б. Ам. метеорол. Соц., 96, 1311–1332, https://doi.org/10.1175/BAMS-D-12-00227.1, 2015.
Икбал, М.: Введение в солнечное излучение, Acadamec Press, Онтарио, 1983.
Яницка Л., Стахлевская И.С., Веселовский И. и Баарс Х.: Temporal
изменение оптических и микрофизических свойств
аэрозоль горения минеральной пыли и биомассы, полученный с помощью дневного рамановского лидара
наблюдения над Варшавой, Польша, Атмос. Окружающая среда., 169, 162–174, https://doi.org/10.1016/j.atmosenv.2017.09.022,
2017.
Джуда-Резлер, К., Рейзер, М., и Удине, Дж. П.: Определение и анализ Распределение источников PM10 во время эпизодов загрязнение воздуха в городских районах Центральной и Восточной Европы: пример зима 2006, Атмос. Окружающая среда, 45, 6557–6566, https://doi.org/10.1016/j.atmosenv.2011.08.020, 2011.
Джуда-Резлер К., Рейзер М., Гузар П., Крюгер Б., Занис П., Сыраков, Д., Катрагкоу Э., Трапп В., Мелас Д., Червенков Х., Тегулиас И. и Галенко Т.: Моделирование эффектов изменение климата на качество воздуха над центральной и Восточная Европа: концепция, оценка и прогнозы, Клим. Рез., 53, 179–203, https://doi.org/10.3354/cr01072, 2012.
Юнг, Э., Альбрехт, Б.А., Фейнгольд, Г., Йонссон, Х.Х., Чуанг, П., и Донахер, С.Л.: Аэрозоли, облака и осадки в Северной Атлантике, наблюдаемые во время эксперимента с аэрозольными облаками на Барбадосе — Часть 1: Распределение и изменчивость, Atmos. хим. Phys., 16, 8643–8666, https://doi.org/10.5194/acp-16-8643-2016, 2016.
Кауфман Ю. Дж., Танре Д. и Буше О.: Аэрозольный снимок со спутника в климатической системе, Природа, 419, 215–223, https://doi.org/10.1038/nature01091, 2002.
Киплинг, З., Стир, П., Джонсон, К.Э., Манн, Г.В., Беллоуэн, Н., Бауэр, С.Е., Бергман, Т., Чин, М., Дил, Т., Ган, С.Дж., Иверсен, Т., Киркевог, А., Коккола, Х., Лю, Х., Луо, Г., ван Нойе, Т., Прингл, К.Дж., фон Зальцен, К., Шульц М., Селанд О., Скейе Р. Б., Такемура Т., Цигаридис К. и Чжан К.: Что контролирует вертикальное распределение аэрозоля? Взаимосвязь между чувствительностью процесса в HadGEM3–UKCA и межмодельным отклонением от AeroCom Phase II, Atmos. хим. Phys., 16, 2221–2241, https://doi.org/10.5194/acp-16-2221-2016, 2016.
Коффи, Б., Шульц, М., Бреон, Ф.М., Дентенер, Ф., Стенсен, Б.М.,
Грисфеллер Дж., Винкер Д., Балкански Ю., Бауэр С. Э.,
Беллоуэн Н., Бернтсен Т., Биан Х.С., Чин М., Диль Т., Истер Р.,
Ган С., Хауглустейн Д. А., Иверсен Т., Киркеваг А.,
Лю Х. Х., Ломанн У., Мире Г., Раш П., Селанд О., Скейе Р. Б.,
Стинрод, С. Д., Стир, П., Тэкетт, Дж., Такемура, Т.,
Цигаридис, К., Вуоло, М. Р., Юн, Дж., и Чжан, К.: Оценка
вертикальное распределение аэрозоля в глобальных моделях аэрозолей
посредством сравнения с измерениями CALIOP: результаты фазы II AeroCom, J.
Геофиз. рез.-атмосфер., 121, 7254–7283,
https://doi.org/10.1002/2015JD024639, 2016.
Лелиевельд Дж., Эванс Дж. С., Фнайс М., Джаннадаки Д. и Поззер А.: вклад источников загрязнения атмосферного воздуха в преждевременная смертность в глобальном масштабе, Nature, 525, 367–371, https://doi.org/10.1038/nature15371, 2015.
Ли, X., Ма, Ю., Ван, Ю., Лю, Н., и Хун, Ю.: Временные и пространственные анализы твердых частиц (PM 10 и PM 2.5 ) и его связь с метеорологическими параметрами над городским городом в Северо-Восточный Китай, Атмос. Рез., 198, 185–193, https://doi.org/10.1016/j.atmosres.2017.08.023, 2017.
Лисок Дж. , Розвадовска А., Педерсен Дж. Г., Маркович К. М., Риттер К., Камински Дж. В., Стружевска Дж., Маццола М., Удисти Р., Бекальи С. и Горецка И.: Радиационное воздействие экстремального горения арктической биомассы, Atmos. хим. Phys., 18, 8829–8848, https://doi.org/10.5194/acp-18-8829-2018, 2018.
Лю, Ю., Франклин, М., Кан, Р., и Кутракис, П. .: Использование аэрозольной оптики толщина для прогнозирования PM 9 на уровне земли1237 2,5 концентрации в районе Сент-Луиса: сравнение между MISR и MODIS, Дистанционный датчик окружающей среды, 107, 33–44, https://doi.org/10.1016/j.rse.2006.05.022, 2007.
Лолли, С. и Ди Джироламо, П.: Подход к анализу основных компонентов для оценки характеристик прибора при разработке рентабельного надежного прибора Сеть атмосферных измерений, J. Atmos. Океан. Тех., 32, 1642–1649, https://doi.org/10.1175/JTECH-D-15-0085.1
Лолли, С., Мадонна, Ф., Росольди, М., Кэмпбелл, Дж. Р., Велтон, Э. Дж., Льюис, Дж. Р., Гу, Ю. и Паппалардо Г.: Влияние различных лидарных измерений и методов обработки данных на оценку прямых радиационных эффектов перистых облаков и аэрозолей, Atmos. Изм. Тех., 11, 1639–1651, https://doi.org/10.5194/amt-11-1639-2018, 2018.
Марину Э., Амиридис В., Биниетоглу И., Цикердекис А., Соломос С., Проетакис Э., Конста Д., Папагианнопулос Н., Цекери А., Властоу Г., Занис П., Балис Д., Вандингер У. и Ансманн А.: Трехмерная эволюция переноса пыли из Сахары в Европу на основе 9-летнего оптимизированного для EARLINET набора данных CALIPSO, Atmos. хим. Phys., 17, 5893–5919, https://doi.org/10.5194/acp-17-5893-2017, 2017.
Маркович К., Чилински М. Т., Лисок Ю., Завадская О. Стахлевская И. С., Яницкая Л., Розвадовская А., Макуч П., Пакшис П., Зелински Т., Петельски Т., Посиняк М., Петручук А., Шкоп, А., и Вестфаль, Д.Л.: Исследование аэрозоля. оптические свойства при дальнем переносе горящей биомассы из Канада в Центральную Европу в июле 2013 г., J. Aeros. Sci., 101, 156–173, https://doi.org/10.1016/j.jaerosci.2016.08.006, 2016.
Масонис С.Дж., Андерсон Т.Л., Коверт Д.С., Капустин В., Кларк А.Д.,
Хауэлл, С., и Мур, К.: Исследование
отношение поглощения к обратному рассеянию морского аэрозоля на береговой линии
исследование аэрозолей в окружающей среде, J. Atmos. Океан. Тех.,
20, 1388–1402, https://doi.org/10.1175/1520-0426(2003)020<1388:ASOTER>2.0.CO;2, 2003. Портнер, Х.-О., Робертс, Д., Скеа, Дж., Шукла, П.Р., Пирани, А., Муфума-Окиа, В., Пеан, К.,
Пидкок Р., Коннорс С., Мэтьюз Дж. Б. Р., Чен Ю., Чжоу X., Гомис М. И.,
Лонной Э., Мэйкок Т., Тигнор М. и Уотерфилд Т.:
Глобальное потепление на 1,5 ∘ C, Специальный отчет МГЭИК о
последствия глобального потепления на 1,5 91 185 ∘ 91 186 C выше доиндустриального уровня и соответствующие глобальные пути выбросов парниковых газов, в
контексте усиления глобального реагирования на
угроза изменения климата, устойчивое развитие и усилия по искоренению
бедность, МГЭИК, 2018 г.
Маттиас В., Балис Д., Бёзенберг Дж., Эйксманн Р., Ярлори М.,
Комгуем Л., Маттис И., Папаяннис А., Паппалардо Г.,
и Перроне, М.: Вертикальное распределение аэрозолей над Европой: Статистические данные.
анализ данных рамановского лидара из 10 европейских
Станции лидарной сети для исследования аэрозолей (EARLINET), J. Geophys. Рез., 109, Д18201,
https://doi.org/10.1029/2004JD004638,
2004.
Маттис И., Ансманн А., Мюллер Д., Вандингер У. и Альтхаузен Д.: Многолетние наблюдения за аэрозолями на двух длинах волн Рамановский лидар в рамках EARLINET, J. Geophys. Рез., 109, Д13203, https://doi.org/10.1029/2004JD004600, 2004.
Мона, Л., Амодео, А., Д’Амико, Г., Джунта, А., Мадонна, Ф., и Паппалардо, Г.: Multi Рамановские лидарные наблюдения за вулканическим облаком Эйяфьятлайокудль над Потенцей, южная Италия, Атмос. хим. физ., 12, 2229–2244, https://doi.org/10.5194/acp-12-2229-2012, 2012.
Мюллер Д., Ансманн А., Маттис И., Теше М., Вандингер У., Альтхаузен, Д., и Пизани, Г.: Лидар, зависящий от типа аэрозоля отношения, наблюдаемые с помощью рамановского лидара, J. Geophys. Рез., 112, Д16202, https://doi.org/10.1029/2006JD008292, 2007.
Навас-Гусман, Ф., Мартуччи, Г., Коллауд Коэн, М., Гранадос-Муньос, М. Дж., Эрво, М., Сикард, М., и Хефеле, А.: Характеристика гигроскопичности аэрозолей с использованием измерений рамановского лидара на станции EARLINET в Пайерне, Атмос. хим. физ., 19, 11651–11668, https://doi.org/10.5194/acp-19-11651-2019, 2019.
Немук, А., Василеску, Дж., Талиану, К., Белеганте, Л., и Николае, D.: Оценка массовых концентраций аэрозоля по измеренному коэффициенту линейной деполяризации частиц (с вертикальным разрешением) и моделированию, Atmos. Изм. Tech., 6, 3243–3255, https://doi.org/10.5194/amt-6-3243-2013, 2013.
Немуц А., Стахлевска И.С., Валилеску Ю., Горска А., Николае, Д., и Талиану, К.: Оптические свойства дальнего действия. Транспортировка вулканического пепла над Румынией и Польшей во время Эйяфьядлайокудля Извержение 2010 г., Acta Geophys., 62, 350–366. https://doi.org/10.2478/s11600-013-0180-7, 2014 г.
Николае Д., Василеску Дж., Талиану К., Биниетоглу И., Николае В., Андрей С. и Антонеску Б.: Алгоритм нейросетевого типирования аэрозолей на основе данных лидара, Атмос. хим. Phys., 18, 14511–14537, https://doi.org/10.5194/acp-18-14511-2018, 2018. Бенавент-Ольтра, Дж. А., Бёкманн, К., Самарас, С., Стахлевска, И. С., Яницка, Л., Баарс, Х., Больманн, С., и Аладос-Арболедас, Л.: Микрофизическая характеристика переносимых на большие расстояния частицы сжигания биомассы из Северной Америки на трех станциях EARLINET, Atmos. хим. физ., 17, 5931–5946, https://doi.org/10.5194/acp-17-5931-2017, 2017.
Пан, X., Чин, М., Гаутам, Р., Биан, Х., Ким, Д. , Коларко, П.Р., Диль, Т.Л., Такемура, Т., Поццоли, Л., Цигаридис, К., Бауэр, С., и Беллоуэн, Н.: Мультимодельная оценка аэрозолей над Южной Азией: общие проблемы и возможные причины, Атмос. хим. Phys., 15, 5903–5928, https://doi.org/10.5194/acp-15-5903-2015, 2015. В., Баарс Х., Биниетоглу И., Бортоли Д., Д’Амико Г., Джунта А., Герреро-Раскадо Дж. Л., Шварц А., Перейра С., Спинелли Н. , Wandinger, U., Wang, X. и Pappalardo, G.: Климатологическая продукция CALIPSO: оценка и предложения от EARLINET, Atmos. хим. Phys., 16, 2341–2357, https://doi.org/10.5194/acp-16-2341-2016, 2016.
Папагианнопулос, Н., Мона, Л., Амодео, А., Д’Амико, Г., Гума Кларамунт, П. , Паппалардо, Г., Аладос-Арболедас , Л., Герреро-Раскадо, Дж. Л., Амиридис, В., Коккалис, П., Апитулей, А., Баарс, Х., Шварц, А., Вандингер, У., Биниетоглу, И., Николае, Д., Бортоли Д., Комерон А., Родригес-Гомес А., Сикард М., Папаяннис А. и Вигнер М.: Метод автоматического типирования аэрозолей на основе наблюдений для EARLINET, Atmos. хим. Phys., 18, 15879–15901, https://doi.org/10.5194/acp-18-15879-2018, 2018.
Папаяннис А., Амиридис В., Мона Л., Цакнакис Г., Балис Д., Бозенберг, Дж., Чайковский А., Де Томази Ф., Григоров И., Маттис И., Митев В., Мюллер Д., Никович С., Перес К., Петручук А., Пизани Г., Раветта Ф., Ризи В., Сикард М., Трикл Т., Вигнер М., Гердинг М., Мамури Р. Э., Д’Амико Г. и Паппалардо, Г.: Систематические лидарные наблюдения Пыль Сахары над Европой в кадре EARLINET (2000–2002 гг.), J. Geophys. Рез., 113, D10204, https://doi.org/10.1029/2007JD009028, 2008.
Паппалардо, Г., Амодео, А., Апитулей, А., Комерон, А., Фройденталер, В., Линне, Х., Ансманн, А. , Босенберг, Дж., Д’Амико Г., Мэттис И., Мона Л., Вандингер У., Амиридис В., Аладос-Арболедас Л., Николае Д. и Вигнер М.: EARLINET: на пути к передовому устойчивому европейскому аэрозолю лидарная сеть, Atmos. Изм. Tech., 7, 2389–2409, https://doi.org/10.5194/amt-7-2389-2014, 2014.
Perrone, M.R., De Tomasi, F., and Gobbi, G.P.: Свойства аэрозоля с вертикальным разрешением по многоволновым лидарным измерениям, Атмос. хим. Phys., 14, 1185–1204, https://doi.org/10.5194/acp-14-1185-2014, 2014.
Petters, M.D., Carrico, C.M., Kreidenweis, S.M., Prenni, A.J., DeMott, П.Дж., Коллетт, Дж.Л., и Моосмюллер, Х.: Облако активность зародышеобразования конденсации аэрозоля горения биомассы, J. Geophys. Рез., 114, D22205, https://doi.org/10.1029/2009JD012353, 2009.
Поуп, С. Б.: Бурные потоки. Издательство Кембриджского университета, Великобритания, 771 стр., 2000. К., Виктори С., Дюкос Ф., Торрес Б., Делегов К., Чоэль М., Пухоль-Сёне Н. и Пьетрас К.: Описание и применение мобильной системы, работающей на — дистанционное зондирование дорожных аэрозолей и измерения на месте, Атмос. Изм. Тех., 11, 4671–4691, https://doi.org/10.5194/amt-11-4671-2018, 2018.
Посфаи М., Геленсер А., Симоникс Р., Арато К., Ли Дж., Хоббс, П.В., и Бусек, П.Р.: Атмосферные смоляные шарики: частицы. от сжигания биомассы и биотоплива, J. Geophys. Рез., 109, Д06213, https://doi.org/10.1029/2003JD004169, 2004.
Проетакис, Э., Амиридис, В., Марину, Э., Биниетоглу, И., Ансманн, А., Вандингер, У., Хофер, Дж. , Йоркс Дж., Новотник Э., Махмудов А., Папаяннис А., Петручук А., Гиалитаки А., Апитулей А., Шкоп А., Муньос Поркар К., Бортоли Д. ., Диониси Д., Альтхаузен Д., Мамали Д., Балис Д., Николае Д., Тетони Э., Либерти Г.Л., Баарс Х., Маттис И., Стахлевска И.С., Вудури К.А., Мона Л., Милонаки М., Перроне М.Р., Коста М.Дж., Сикард М., Папагианнопулос Н., Сиомос Н., Бурлицци П., Поли Р., Энгельманн Р. ., Абдуллаев С. и Паппалардо Г.: Оценка EARLINET произведения коэффициента обратного аэрозольного рассеяния CATS Level 2, Atmos. хим. физ., 19, 11743–11764, https://doi.org/10.5194/acp-19-11743-2019, 2019.
Цинь, К., Цзоу, Дж., Го, Дж., Лу, М., Билал, М. ., Чжан, К., Ма, Ф., и Чжан, Y.: Оценка концентраций PM1 по MODIS за Дельта реки Янцзы в Китае в 2014–2017 гг. Атмос. Окружающая, 195, с. 149–158, https://doi.org/10.1016/j.atmosenv.2018.09.054, 2018.
Reizer, M. and Juda-Rezler, K.: Объяснение высоких концентраций PM 10 наблюдается в польских городских районах, Air Qual. Атмос Здоровье., 9, 517–531, https://doi.org/10.1007/s11869-015-0358-z, 2015.
Рост Дж., Холст Т., Сен Э., Клингнер М., Анке К., Аренс Д. и Майер, Х.: Изменчивость концентраций PM10 зависимости от метеоусловий, междунар. Дж. Окружающая среда. Опрос., 36, 3–18, https://doi.org/10.1504/IJEP.2009.021813, 2009.
Сакаи Т., Нагаи Т., Зайзен Ю. и Мано Ю.: Линейное обратное рассеяние
измерения коэффициента деполяризации минералов, морской соли,
и частицы сульфата аммония, смоделированные в лабораторной камере, Appl.
Опт., 49, 4441–4449, https://doi.org/10.1364/AO.49.004441, 2010.
Шаап, М., Апитулей, А., Тиммерманс, Р. глубина и PM2,5 в Кабау, Нидерланды, Атмос. хим. Phys., 9, 909–925, https://doi.org/10.5194/acp-9-909-2009, 2009.
Шефер, К., Эмейс, С., Хоффманн, Х., и Ян, С. .: Влияние смешения высота слоя при загрязнении атмосферного воздуха в городских и пригородных районах // Метеорология. З., 15, 647–658, https://doi.org/10.1127/0941-2948/2006/0164, 2006.
Шмайссер, Л., Эндрюс, Э., Огрен, Дж. А., Шеридан, П., Джефферсон, А., Шарма, С., Ким, Дж. Э., Шерман, Дж. П., Соррибас М., Калапов И., Арсов Т., Ангелов К., Майоль-Брасеро О.Л., Лабушань К., Ким С.-В., Хоффер А., Лин Н.-Х. , Chia, H.-P., Bergin, M., Sun, J., Liu, P. и Wu, H.: Классификация типов аэрозолей с использованием оптических свойств поверхностных спектральных аэрозолей in situ, Atmos. хим. Phys., 17, 12097–12120, https://doi.org/10.5194/acp-17-12097-2017, 2017.
Сайнфелд, Дж. Х., Бретертон, К., Карслоу, К. С., Коу, Х., ДеМотт, П. Дж.,
Данли, Э. Дж., Фейнгольд, Г. , Ган, С., Гюнтер, А. Б.,
Кан Р., Краукунас И., Крайденвайс С. М., Молина М. Дж., Ненес А.,
Пеннер, Дж. Э., Пратер, К. А., Раманатан, В.,
Рамасвами В., Раш П.Дж., Равишанкара А.Р., Розенфельд Д., Стивенс,
Г. и Вуд Р.: Улучшение нашего фундаментального
понимание роли аэрозольно-облачных взаимодействий в климате
система, П. Натл. акад. науч. США, 113, 5781–5790,
https://doi.org/10.1073/pnas.1514043113, 2016 г.
Сикард, М., Рокаденбош, Ф., Реба, М. Н. М., Комерон, А., Томас, С., Гарсия-Вискайно, Д., Батет, О., Барриос, Р., Кумар, Д., и Балдасано , JM: Сезонная изменчивость оптических свойств аэрозолей, наблюдаемая с помощью рамановского лидара на площадке EARLINET над северо-востоком Испании, Atmos. хим. Phys., 11, 175–190, https://doi.org/10.5194/acp-11-175-2011, 2011.
Сиомос Н., Балис Д.С., Пупкоу А., Лиора Н., Димопулос С., Мелас Д., Джаннакаки Э., Филиоглу М., Басарт С. и Чайковский А.: Исследование качества смоделированных профилей аэрозоля на основе объединенных данных лидара и солнечного фотометра, Атмос. хим. Phys., 17, 7003–7023, https://doi.org/10.5194/acp-17-7003-2017, 2017.
Сиомос, Н., Балис, Д.С., Вудури, К.А., Гианнакаки, Э., Филиоглу, М., Амиридис, В., Папаяннис, А., и Фрагкос, К.: Согласованы ли климатологии EARLINET и AERONET? Дело Салоники, Греция, Атмос. хим. Phys., 18, 11885–11903, https://doi.org/10.5194/acp-18-11885-2018, 2018.
Шарма А., Мандал Т., Шарма С., Шукла Д. , и Сингх, С.: Отношения приземного озона с его прекурсорами, твердыми частицами материя и метеорология над Дели, J. Atmos. хим., 74, 451–474, https://doi.org/10.1007/s10874-016-9351-7, 2017.
Стахлевска И., Пёндловски М., Мигач С., Шкоп А., Зелинска, А. и Свачина П.: Облакомерные наблюдения границы слой над Варшавой, Польша, Acta Geophys., 60, 1386–1412, https://doi.org/10.2478/s11600-012-0054-4, 2012.
Стахлевская И.С., Коста-Сурос М. и Альтхаузен Д.: Рамановский лидар
профилирование водяного пара над Варшавой, Польша, Atmos.
рез., 194, 258–267, https://doi.org/10.1016/j. atmosres.2017.05.004, 2017a.
Стахлевская И.С., Завадска О. и Энгельманн Р.: Влияние аномальной жары условия на оптические свойства аэрозоля, полученные от спутникового и наземного дистанционного зондирования над Польшей, Дистанционное зондирование, 9, 1199, https://doi.org/10.3390/rs
99, 2017b.Стахлевская И.С., Самсон М., Завадска О., Харенда К.М., Яницка Л., Почта П., Щепаник Д., Хиз Б., Ван Д., и Борек, К. Тетони, Э., Проетакис, Э., Сиомос, Н., Немуц, А., Хойницкий, Б. Х., Маркович К. М., Петручук А., Шкоп, А., Альтхаузен, Д., Стебель, К., Шюттемейер, Д., и Зенер, К.: Изменение свойств местных городских аэрозолей за счет дальний перенос горящего аэрозоля биомассы, Remote Sens., 10, 412, https://doi.org/10.3390/rs10030412, 2018.
Штейн, А. Ф., Дракслер, Р. Р., Рольф, Г. Д., Стандер, Б. Дж. Б., Коэн, М. Д.,
и Нган, Ф.: HYSPLIT NOAA Atmospheric.
Система моделирования переноса и дисперсии, B. Am. метеорол. Соц., 96,
2059–2077, https://doi.org/10.1175/BAMS-D-14-00110. 1, 2015.
Стокер Т., Цинь Д., Платтнер Г., Тигнор М., Аллен С., Бошунг Дж., Науэльс А., Ся Ю., Бекс В. и Мидгли П.: МГЭИК, 2013: Изменение климата 2013: Основы физических наук, Вклад Рабочая группа I по пятой оценке Отчет Межправительственной группы экспертов по изменению климата, Кембриджский университет. Press, Кембридж, Великобритания, и Нью-Йорк, 1535 стр., 2013 г.
Стулл, Р. Б.: Введение в метеорологию пограничного слоя, Kluwer Academic Publishers, Дордрехт, Нидерланды, 1988.
Щепаник, Д. и Маркович, К.: Соотношение между оптические свойства аэрозоля на заднем плане окружающая среда, Атмос. Опрос. рез., 9, 246–256, https://doi.org/10.1016/j.apr.2017.10.001, 2018.
Щепаник Д., Тетони Э., Ван Д. и Стахлевска И.: на основе лидара Отделение загрязненной пыли над Варшавой (кейс Учеба 09август 2013 г.) 29-й Международный лазерный радар Конференция, Хэфэй, Китай, 24–28 июня 2019 г., 1–5 июня 2019 г.
Шкоп А. и Петручук А.: Анализ переноса аэрозолей над южными
Польша в августе 2015 г. на основе синергии
методы дистанционного зондирования и обратной траектории, J. Appl. Удаленный. сен.,
11, 016039, https://doi.org/10.1117/1.JRS.11.016039, 2017.
Тан, И. Н.: Химическое и размерное воздействие гигроскопического аэрозоля на свет коэффициенты рассеяния, J. Geophys. Рез., 101, 19245–19250, https://doi.org/10.1029/96JD03003, 1996.
Издательская группа EARLINET 2000–2015: EARLINET All 2000–2015, World Data Центр климата (WDCC) в DKRZ, https://doi.org/10.1594/WDCC/EARLINET_All_2000-2015, 2018.
Тянь, П., Цао, X., Чжан, Л., Сун, Н., Сун, Л., Логан, Т. ., Ши Дж., Ван Ю., Цзи Ю., Линь Ю., Хуан З., Чжоу Т., Ши Ю. и Чжан Р.: Вертикальное распределение аэрозолей и оптические свойства над Китай от долговременного спутникового и наземного ДЗЗ, Атмос. хим. Phys., 17, 2509–2523, https://doi.org/10.5194/acp-17-2509-2017, 2017.
Трикл, Т., Фогельманн, Х., Флентье, Х. и Рис, Л.: Стратосферный озон в северных широтах шлейфы огня — сезон дыма 2013 г. над Центральной Европой, Атмос. хим. Phys., 15, 9631–9649, https://doi.org/10.5194/acp-15-9631-2015, 2015.
Trippetta, S., Sabia, S., and Caggiano, R.: Мелкие аэрозольные частицы (PM 1 ): Природный и антропогенный вклад и риск для здоровья оценка, Air Qual. Атмос. Hlth., 9, 621–629, https://doi.org/10.1007/s11869-015-0373-0, 2016.
Веселовский И., Колготин А., Грязнов В., Мюллер Д., Вандингер У., и Уайтмен, Д. Н.: Инверсия с регуляризацией для получение параметров тропосферного аэрозоля из многоволнового лидара звучание, прил. Opt., 41, 3685–3699, https://doi.org/10.1364/AO.41.003685, 2002.
Валашек К., Крыза М. и Вернер М.: Роль выбросов прекурсоров о концентрации приземного озона во время летний сезон в Польше, J. Atmos. хим., 75, 181–204, https://doi.org/10.1007/s10874-017-9371-y, 2018.
Wandinger, U., Freudenthaler, V., Baars, H., Amodeo, A., Engelmann, R., Mattis, I., Groß, S., Pappalardo, G., Giunta, А., Д’Амико Г., Чайковский А., Осипенко Ф., Слесар А., Николае Д. , Белеганте Л., Талиану К., Сериков И., Линне Х., Янсен Ф., Апитулей А., Уилсон К.М., де Грааф М., Трикл Т., Гиль Х., Адам М., Комерон А., Муньос-Поркар К., Рокаденбош Ф. ., Сикард М., Томас С., Ланге Д., Кумар Д., Пухадас М., Молеро Ф., Фернандес А. Х., Аладос-Арболедас Л., Браво-Аранда Дж. А., Навас -Гусман Ф., Герреро-Раскадо Х.Л., Гранадос-Муньос М.Дж., Прейслер Дж., Вагнер Ф., Гауса М., Григоров И., Стоянов Д., Ярлори М., Ризи, В., Спинелли, Н., Боселли, А., Ван, X., Ло Федо, Т., Перроне, М. Р., Де Томази, Ф. и Бурлицци, П.: Кампании по взаимному сравнению приборов EARLINET: обзор стратегии и результаты , Атмос. Изм. Тех., 9, 1001–1023, https://doi.org/10.5194/amt-9-1001-2016, 2016.
Ван, Д., Стахлевска, И. С., Сонг, X., Хиз, Б. и Немук, А.: Изменчивость пограничного слоя над городской континентальный участок на основе 10 лет активного дистанционного зондирования наблюдений в Варшаве, Remote Sens., обзор, 2019 г.
Ван Дж. и Кристофер С.А.: Сравнение спутников
оптическая толщина аэрозоля и масса PM 2,5 :
Последствия для изучения качества воздуха, Geophys. Рез. Письма, 30, 2095,
https://doi.org/10.1029/2003GL018174, 2003.
Винкер, Д. М., Хант, У. Х., и Макгилл, М. Дж.: Начальное исполнение оценка КАЛИОП, Геофиз. Рез. Летт., 34, Л19803, https://doi.org/10.1029/2007GL030135, 2007 г.
Вольф, Х. и Перри, Л.: Мониторинг политики: Тенденции в законодательстве о чистом воздухе в Европа: Твердые частицы и зоны с низким уровнем выбросов, Rev. Окружающая среда. Экон. Policy, 4, 293–308, https://doi.org/10.1093/reep/req008, 2010.
Се, К., Нисидзава, Т., Сугимото, Н., Мацуи, И., и Ван, З. .: Характеристики оптических свойств аэрозолей в условиях загрязнения и Эпизоды азиатской пыли над Пекином, Китай, Прил. Опт., 47, 4945–4951, https://doi.org/10.1364/AO.47.004945, 2008.
Zang, Z.L., Wang, W.Q., You, W., Li, Y., Ye, F., and Wang, C.M.:
Оценка приземных концентраций PM 2,5 в
Пекин, Китай с использованием оптической толщины аэрозоля и параметров температуры
инверсионный слой, наук. Total Environ., 575, 1219–1227, https://doi. org/10.1016/j.scitotenv.2016.09.186, 2017.
Завадска О., Маркович К., Петручук А., Зелински Т. ., и Ярославский, J.: Воздействие городского загрязнения, выбрасываемого в Варшава по аэрозольным свойствам, Atmos. Окружающая, 69, 15–28, https://doi.org/10.1016/j.atmosenv.2012.11.065, 2013.
Zhang, H., Wang, Y., Hu, J., Ying, Q. и Hu, X.-M. : Отношения между метеорологические параметры и критерии загрязнителей атмосферного воздуха в трех мегаполисах Китая, Environ. рез., 140, 242–254, https://doi.org/10.1016/j.envres.2015.04.004, 2015.
Чжэн, С., Поззер, А., Цао, С. X., и Леливельд, Дж.: Долгосрочные (2001–2012 гг.) концентрации мелких твердых частиц (PM 2,5 ) и воздействие на здоровье человека в Пекине, Китай, атмосфер. хим. Phys., 15, 5715–5725, https://doi.org/10.5194/acp-15-5715-2015, 2015.
Паттернирование эпителия желудочно-кишечного тракта для придания регионарно-специфических функций
- Список журналов
- Рукописи авторов HHS
- PMC6615902
Дев Биол. Авторская рукопись; доступно в PMC 20199 июля.
Опубликовано в окончательной редакции как:
Dev Biol. 2018 15 марта; 435(2): 97–108.
Опубликовано онлайн 2018 г., 12 января. DOI: 10.1016/j.ydbio.2018.01.006
PMCID: PMC6615902
NIHMSID: NIHMS1529689
PMID: 2939095
999689PMID: 2939095
(GSIRILIN тракта, проще говоря, можно описать как выстланную эпителием мышечную трубку, проходящую вдоль цефалокаудальной оси от ротовой полости до анального отверстия. Хотя общая архитектура органов желудочно-кишечного тракта сохраняется от начала до конца, наличие различных структур эпителиальной ткани и уникальных типов эпителиальных клеток в каждом органе позволяет каждому выполнять различные пищеварительные функции, необходимые для эффективного усвоения питательных веществ. Пространственно-временная регуляция сигнальных путей и нижестоящих факторов транскрипции контролирует морфогенез эпителия ЖКТ во время развития, чтобы обеспечить существенные регионально-специфические эпителиальные структуры и функции. Здесь мы обсуждаем основные функции каждого органа желудочно-кишечного тракта и резюмируем разнообразие эпителиальных структур, присутствующих вдоль цефалокаудальной оси желудочно-кишечного тракта. Далее мы обсудим результаты, прежде всего полученные на генетических моделях мышей, которые определили роль ключевых факторов транскрипции во время эпителиального морфогенеза, включая p63, SOX2, SOX15, GATA4, GATA6, HNF4A и HNF4G. Кроме того, мы изучаем, как сигнальные пути Hedgehog, WNT и BMP способствуют определению уникальных особенностей эпителия вдоль цефалокаудальной оси желудочно-кишечного тракта. Наконец, мы исследуем молекулярные механизмы, контролирующие региональную цитодифференцировку органоспецифических типов эпителиальных клеток в желудочно-кишечном тракте, концентрируясь на желудке и тонкой кишке. Выявление механизмов формирования эпителиального паттерна желудочно-кишечного тракта у мышей предоставило фундаментальные знания для разработки и уточнения трехмерных моделей органотипических культур желудочно-кишечного тракта, таких как модели, полученные в результате направленной дифференцировки плюрипотентных стволовых клеток человека, и те, которые получены непосредственно из образцов тканей человека. Дальнейшее изучение этих путей, несомненно, даст жизненно важное представление о механизмах развития и заболеваний ЖКТ и может открыть новые возможности для инновационной тканевой инженерии и подходов персонализированной медицины к лечению желудочно-кишечных заболеваний.
Ключевые слова: эпителий желудочно-кишечного тракта, развитие желудочно-кишечного тракта, регионализация, морфогенез, цитодифференцировка, факторы транскрипции, сигнальные пути
Желудочно-кишечный тракт (ЖКТ) представляет собой выстланную эпителием мышечную трубку, простирающуюся от ротовой полости до заднего прохода. Органы желудочно-кишечного тракта, включая пищевод, желудок, тонкую кишку и толстую кишку, функционируют в координации с поджелудочной железой, печенью и желчным пузырем, выполняя жизненно важные задачи пищеварения и всасывания. Желудочно-кишечный тракт также содержит самый большой в организме резервуар лимфоидной ткани и кишечную нервную систему с большим количеством нейронов, чем в спинном мозге. Общая архитектура органов желудочно-кишечного тракта сохраняется со всеми эмбриональными зародышевыми слоями, вносящими свой вклад в ткани каждого органа (12). Самый внутренний слой слизистой оболочки состоит из эпителия, происходящего из энтодермы. Эпителий поддерживается собственной пластинкой, слоем соединительной ткани, и мышечной слизистой оболочкой, тонким слоем гладких мышц, происходящими из мезодермы. Под слизистой оболочкой находится подслизистая оболочка, содержащая рыхлую коллагеновую ткань мезодермы, жировую ткань и крупные кровеносные и лимфатические сосуды. Эктодермальные кишечные ганглии также находятся в подслизистой оболочке. Muscleis externa, толстый слой мезодермальных мышц, содержащий дополнительные энтеральные ганглии, оборачивается вокруг слизистого и подслизистого слоев и генерирует перистальтические силы, которые перемещают содержимое по кишечнику. Обычно наружная мышечная оболочка состоит из гладких мышц; однако пищевод и анальный сфинктер содержат скелетные мышцы. Наконец, желудочно-кишечный тракт покрыт адвентицией или серозой, происходящей из мезодермы.
Открыть в отдельном окне
Все эмбриональные зародышевые листки входят в состав тканевых слоев органов ЖКТ. Энтодерма (оранжевая) разделена на три области и дает начало самому внутреннему эпителиальному слою всех органов ЖКТ. Многослойный плоский эпителий пищевода (Е) и преджелудка (FS), а также простой цилиндрический эпителий заднего отдела желудка (HS) и проксимального отдела двенадцатиперстной кишки (D) тонкой кишки происходят из энтодермы передней кишки. Соединение между многослойным плоским эпителием и простым столбчатым эпителием называется плоскостолбчатым соединением (SCJ). Энтодерма средней кишки дает начало простому столбчатому эпителию дистального отдела двенадцатиперстной кишки, тощей (J) и подвздошной (I) тонкой кишки, а также простому столбчатому эпителию проксимального отдела толстой кишки (PC). Простой цилиндрический эпителий дистального отдела толстой кишки (DC) возникает из энтодермы задней кишки. Под эпителием (Ep), собственной пластинкой (LP), мышечной слизистой оболочкой (MM), подслизистой оболочкой (SM), сосудами, лимфатическими сосудами и внешней мышечной оболочкой (CM, круговая мышца; LM, продольная мышца) слои развиваются из мезодермы (зеленый) . Энтеральная нервная система происходит из эктодермы, а ганглии находятся в подслизистой оболочке и наружной мышечной оболочке (синий). Кишечная трубка покрыта адвентицией из мезодермы (А) или серозной оболочкой (S).
Хотя общая трехмерная структура органов желудочно-кишечного тракта сохраняется, каждый орган и даже определенные области внутри органа выполняют специализированные функции, которые в совокупности приводят к эффективному усвоению питательных веществ. В частности, уникальные особенности эпителия каждого органа обеспечивают основу для региональных специфических функций органов, необходимых для пищеварения и всасывания. Структура эпителия и наличие органоспецифических типов эпителиальных клеток определяют региональные специфические функции вдоль желудочно-кишечного тракта. Цель этого обзора — объяснить, как ключевые факторы транскрипции и сигнальные пути способствуют регионализации эпителия ЖКТ. Исследования с использованием генетически модифицированных моделей мышей были первичным источником информации о механизмах, с помощью которых специфические факторы транскрипции и сигнальные пути управляют регионализацией эпителия ЖКТ во время развития. Совсем недавно исследования с использованием трехмерных культур органоидов in vitro также пролили свет на механизмы регионализации. В первой половине этого обзора будет обсуждаться, как факторы транскрипции и восходящие сигнальные пути формируют различные трехмерные структуры эпителия каждого органа ЖКТ. Во второй половине будет обсуждаться, как факторы транскрипции и восходящие сигнальные пути контролируют специфические программы цитодифференцировки в желудочно-кишечном тракте, уделяя особое внимание эпителию желудка и эпителию кишечника.
A. Разнообразие эпителиальных структур в желудочно-кишечном тракте
Прежде чем обсуждать молекулярные механизмы, лежащие в основе архитектурного паттерна эпителия ЖКТ, мы должны сначала оценить структурные различия, которые существуют между эпителиями желудочно-кишечного тракта, и то, как эпителиальная структура каждого органа ЖКТ напрямую поддерживает его уникальные пищеварительные и всасывающие функции. Пищевод представляет собой мышечную трубку, которая транспортирует проглоченные материалы из ротовой полости в желудок. Его эпителий многослойный, с несколькими слоями клеток, обеспечивающими защиту, необходимую перед лицом сил трения, возникающих при прохождении непереваренных и частично переваренных материалов. У грызунов эпителий пищевода ороговевает и распространяется в проксимальный отдел желудка, образуя желудочную область, известную как преджелудок. Отсутствуя в желудке человека, преджелудок грызунов служит зоной удержания пищи и в некоторой степени отвечает за механическое пищеварение (San Roman and Shivdasani, 2011). Простой столбчатый зарождающийся эпителий пищевода/переднего желудка претерпевает морфогенез во время развития, чтобы дать начало зрелому многослойному эпителию, который состоит из базального слоя пролиферирующих клеток и супрабазального слоя дифференцированных клеток (Zhang et al., 2016).
Имеется отчетливое изменение эпителиальной архитектуры желудочно-кишечного тракта при переходе от пищевода/преджелудка к железистому заднему желудку (). В качестве места химического и механического пищеварения простой столбчатый железистый эпителий желудка идеально подходит для высокоэффективной секреции кислоты и выработки пищеварительных ферментов. Граница между многослойным плоским эпителием пищевода/переднего желудка и простым столбчатым эпителием железистого желудка называется плоскостолбчатым соединением (SCJ) (4). Понимание того, как эти различные эпителиальные структуры образуются и поддерживаются в SCJ, имеет важное значение, поскольку метаплазия происходит в этих тканях при заболеваниях ЖКТ, включая пищевод Барретта, аденокарциному пищевода и аденокарциному желудка.
Тонкий кишечник является местом всасывания питательных веществ, а также основным компонентом врожденной иммунной системы. Существенным для функции тонкой кишки является большая площадь поверхности ее простого столбчатого эпителия, который имеет множество складок с глубокими бороздками, называемыми криптами, и пальцевидными выступами, называемыми ворсинками () (Chin et al. , 2017; Noah et al., 2017; Noah et al. al., 2011; Thompson and Battle, 2014; Walton et al., 2016a). Тонкий кишечник можно разделить на три функциональных отдела: двенадцатиперстную кишку, тощую кишку и подвздошную кишку. Эпителиальные клетки двенадцатиперстной кишки синтезируют пищеварительные ферменты, которые секретируются или локализуются на мембране щеточной каймы. Эти ферменты вместе с желчью, вырабатываемой печенью и ферментами поджелудочной железы, завершают переваривание белков, жиров и углеводов (Binder and Reuben, 2005; Jeejeebhoy, 2002). Эпителий тощей кишки выполняет основную часть поглощения питательных веществ (Binder and Reuben, 2005; Davis and Attie, 2008; Jeejeebhoy, 2002). Эпителий подвздошной кишки отвечает за абсорбцию витамина B12 в дополнение к солям желчных кислот, которые рециркулируются в печени (Binder and Reuben, 2005; Davis and Attie, 2008; Jeejeebhoy, 2002). Понимание того, как правильно регионаризован эпителий тонкой кишки, должно дать фундаментальное представление о синдроме короткой кишки (СКК) и может привести к новым вариантам лечения СКК.
Подобно тонкому кишечнику, эпителий толстого кишечника выполняет функцию всасывания. Выстланные простым столбчатым эпителием, расположенным в виде крипт, но лишенным ворсинок, эти эпителиальные клетки в основном поглощают воду и электролиты (4) (Jeejeebhoy, 2002). Как и тонкий кишечник, толстый кишечник также можно разделить на три отдела: слепую кишку, ободочную кишку и прямую кишку. Хотя эпителий тонкого и толстого кишечника имеет много общего, наличие ворсинок и клеток Панета в тонком кишечнике является одной из отличительных черт. Понимание того, как эпителий этих органов специализирован и регионализован, важно, учитывая, что этот процесс может быть нарушен при заболевании. Например, метаплазия клеток Панета или аномальное присутствие клеток Панета в толстой кишке иногда возникает при воспалительных заболеваниях кишечника и колоректальном раке (Tanaka et al., 2001; Wada et al., 2005).
B. Ключевые факторы транскрипции и сигнальные молекулы, которые контролируют формирование структурного паттерна эпителия желудочно-кишечного тракта
i.
p63, главный регулятор морфогенеза многослойного плоского эпителия. Фактор транскрипции p63 считается главным регулятором развития многослойного плоского эпителия (Danily et al., 2004; Koster et al., 2004; Mills et al., 1999; Romano et al., 2012; Senoo). и др., 2007; Ван и др., 2011; Ян и др., 1999; Ю и др., 2005). р63 необходим для установления многослойного эпителия и, следовательно, обычно отсутствует в столбчатом эпителии. Исследования эмбрионов мышей с нокаутом p63 выявили, что правильный эпителиальный морфогенез в пищеводе и кардиальном отделе желудка требует наличия p63 с его отсутствием, предшествующим этому домену (Wang et al., 2011). В частности, эпителий пищевода или преджелудка, лишенный p63, не может расслаиваться и вместо этого остается столбчатым. Цитокератины столбчато-клеточного типа сохраняются в p63-/- пищеводе и кардиальном отделе желудка вместо цитокератинов многослойно-клеточного типа. Изменения профилей экспрессии цитокератина, вероятно, непосредственно способствуют изменению формы клеток в мутантных тканях. Более того, Хаттон и др. показали, что определенный набор цитокератинов, экспрессируемых в конкретной структуре эпителиальной ткани, необходим и специфичен для структурных и функциональных потребностей этой ткани (Hutton et al., 1998). Например, цитокератин СК18 столбчатого типа не может полностью компенсировать потерю цитокератина СК14 стратифицированного типа в коже с мутацией СК14 (Hutton et al., 1998). Помимо изменений в форме клеток и структуре эпителия, эпителий пищевода и преджелудка с нулевым p63 также демонстрирует признаки слизистой метаплазии, включая наличие бокаловидных клеток кишечного типа и экспрессию кишечных маркеров, таких как Villin, что предполагает потерю p63 как возможный механистический шаг в метаплазии. наблюдается при пищеводе Барретта. Далее подчеркивая решающую роль p63 в развитии многослойного эпителия, эктопической экспрессии p63 в простом эпителии легких достаточно, чтобы преобразовать его в многослойный (Romano et al., 2012).
Открыть в отдельном окне
Экспрессия и функции ключевых транскрипционных факторов, необходимых для регионализации эпителия желудочно-кишечного тракта. Черные столбцы представляют профили и градиенты экспрессии факторов транскрипции вдоль цефалокаудальной оси желудочно-кишечного тракта, включая пищевод (E), преджелудок (FS), задний желудок (HS), двенадцатиперстную кишку (D), тощую кишку (J), подвздошную кишку (I), слепую кишку. (Ce) и толстой кишки (C). В таблице обобщена роль(и) каждого фактора транскрипции в формировании и развитии эпителия желудочно-кишечного тракта.
ii. SOX2 и SOX15, факторы транскрипции, играющие важную роль в регионализации многослойного плоского эпителия.
SOX2, транскрипционный фактор семейства Sry-подобных HMG box, присутствует в изобилии в ранней дорсальной энтодерме передней кишки, когда кишечная трубка разделяется на пищевод и трахею (Que et al., 2007; Sherwood et al., 2009). SOX2 остается высоко экспрессированным в пищеводе во время стратификации эпителия, что указывает на его роль в этом процессе (Que et al., 2007) (3). Так как SOX2 нулевые эмбрионы погибают до гаструляции (Avilion et al. , 2003), роль SOX2 в развитии GI была изучена с использованием гипоморфных мутантов (Que et al., 2007). Эти исследования предоставляют убедительные доказательства того, что для морфогенеза и дифференцировки эпителия пищевода требуется SOX2. В то время как у гипоморфных мутантных эмбрионов SOX2 с наиболее выраженным снижением уровня SOX2 (~5% нормального уровня SOX2) развивается атрезия пищевода (EA) и трахеопищеводный свищ (TEF) из-за дефектов разделения трубок, у эмбрионов, сохраняющих несколько более высокие уровни SOX2 (~18% нормального уровня уровень SOX2) вместо этого имеют дефекты стратификации и развития эпителия пищевода. У этих мутантов эпителий пищевода содержит области аномальных столбчатых клеток, а не многослойных плоскоклеточных клеток. Соответственно уменьшается количество клеток р63+. Более того, некоторые SOX2-дефицитные клетки пищевода экспрессируют маркеры столбчатых эпителиальных клеток желудка и кишечника. Эти исследования указывают на подавление SOX2 как на предполагаемый механизм развития пищевода Барретта и его перехода в аденокарциному (Chen et al. , 2008; Que et al., 2007). Потеря SOX2 была связана с более низкой выживаемостью у пациентов с аденокарциномой пищевода (Honing et al., 2014). И наоборот, активация SOX2 была связана с плоскоклеточным раком (Bass et al., 2009).; Лю и др., 2013).
Экспрессия SOX2 распространяется за пределы эпителия пищевода в преджелудок мышей, который экспрессирует высокие уровни SOX2, и в железистый желудок мыши/человека, который экспрессирует относительно низкие уровни SOX2 по сравнению с пищеводом и преджелудком (Que et al. , 2007). Как и в пищеводе, дефицит SOX2 в развивающемся преджелудке отодвигает эпителий кзади. Мутанты содержат области простого столбчатого эпителия с характеристиками, отражающими характеристики железистого цилиндрического желудка (Que et al., 2007). Удивительным, возможно, является тот факт, что развитие железистого желудка оказывается нормальным у гипоморфных эмбрионов SOX2 (Que et al., 2007). Возможно, что уровень SOX2, сохраняющийся в развивающемся железистом желудке мутантов, достаточен, чтобы направлять правильное развитие эпителия. С появлением моделей на основе стволовых клеток in vitro, которые имитируют развитие желудка в чашке, абсолютная потребность в SOX2 для развития железистого желудка может быть более легко удовлетворена (McCracken et al., 2014; McCracken et al., 2017).
Хотя SOX2 обычно отсутствует в кишечном эпителии, исследования эмбрионов, эктопически экспрессирующих SOX2 в кишечнике, дали информацию о SOX2 как доминирующем драйвере развития эпителия передней кишки. Присутствие SOX2 в развивающемся тонком кишечнике антериоризует эпителий, указывая на то, что SOX2 оказывает доминантное влияние на управление фенотипами эпителия пищевода/желудка (Raghoebir et al., 2012). Экспрессия маркеров типа эпителиальных клеток кишечника снижается, а экспрессия маркеров типа эпителиальных клеток желудка и преджелудка/пищевода стимулируется. Эпителиальная архитектура, хотя и остается столбчатой, аномальна с изменением количества и формы ворсинок. В совокупности эти исследования предполагают, что SOX2 является транскрипционным фактором, играющим существенную роль в определении идентичности многослойного плоского эпителия.
Недавняя работа показала, что SOX15, фактор транскрипции семейства SOX с аминокислотной идентичностью SOX2 на 75%, также контролирует экспрессию генов в клетках многослойного эпителия пищевода человека, регулируя экспрессию значительной части транскриптома клеток многослойного плоского эпителия пищевода ( ) (Sulahian et al., 2015; van de Wetering and Clevers, 1993). Это исследование было выполнено с использованием in vitro делеции SOX15 в модели культуры клеток пищевода, и поэтому будет важно оценить роль SOX15 в регионализации эпителия ЖКТ с использованием мышиных моделей in vivo или моделей органоидов пищевода. Хотя Sox15 нулевых мышей, как сообщалось, были в целом нормальными и фертильными, углубленный анализ развития пищевода в отсутствие SOX15 не проводился (Maruyama et al., 2005). В соответствии с предполагаемой ролью SOX15 как фактора транскрипции стратифицированных эпителиальных клеток, SOX15 отсутствует в столбчатых метапластических поражениях пищевода Барретта (Sulahian et al. , 2015). Удивительно, но хотя SOX15 отсутствует в большинстве аденокарцином пищевода, он остается экспрессированным примерно в 20-30% аденокарцином пищевода (Sulahian et al., 2015). Это говорит о том, что SOX15 может повторно активироваться во время прогрессирования от пищевода Баррета до рака. Как и ожидалось, рак SOX15+ экспрессирует маркеры как стратифицированных плоскоклеточных, так и столбчатых клонов, и будет важно определить патофизиологию таких поражений.
iii. CDX2, основной регулятор развития столбчатого кишечного эпителия.
Транскрипционный фактор CDX2, содержащий гомеобоксный домен, функционирует как основной регулятор развития столбчатого кишечного эпителия (Beck et al., 1999; Chawengsaksophak et al., 1997; Gao et al., 2009; Grainger et al., 2010; Stringer). и др., 2012). Во время развития мышей экспрессия CDX2 становится заметной в развивающейся задней кишке на ст. E8.5, становится ограниченной кишечником на границе передней и средней кишки на ст. E12.5 и сохраняется на этом уровне на протяжении всей взрослой жизни (Beck et al. , 19).95; Бек и др., 1999; Чавенгсаксофак и др., 2004). Проксимальная граница экспрессии CDX2 встречается с дистальной границей экспрессии SOX2 в желудочно-кишечном соединении. Исследования мутантов SOX2 и CDX2 демонстрируют, что установление баланса между SOX2 и CDX2 необходимо для правильного формирования регионального паттерна на этом стыке. Эктопическая экспрессия CDX2 в железистой слизистой оболочке желудка мыши индуцирует экспрессию кишечных генов, и появляются кишечноподобные бокаловидные клетки (Mutoh et al., 2002; Silberg et al., 2002). Совсем недавно Цзян и др. сообщают, что эктопическая экспрессия CDX2 в переходной зоне клеток SCJ индуцирует кишечную метаплазию in vivo, имитируя метаплазию, характерную для пищевода Барретта (Jiang et al., 2017). С др. стороны, эктопическая экспрессия SOX2 в кишечнике ингибирует способность CDX2 связываться со своими генами-мишенями, приводя к фенотипу, подобному передней кишке, или антериоризации кишечника (Raghoebir et al., 2012).
Кроме того, в соответствии с идеей о том, что развивающийся кишечник должен иметь надлежащим образом сбалансированную экспрессию SOX2-CDX2, удаление CDX2 из ранней развивающейся средней кишки посредством условного нокаута индуцирует экспрессию SOX2. Соответственно, CDX2-дефицитная, SOX2-экспрессирующая средняя кишка становится антериоризированной, экспрессируя p63, основной регулятор развития стратифицированного эпителия (Gao et al., 2009).). Совпадая с экспрессией p63 и SOX2, мутантный по CDX2 эпителий кишечника теряет столбчатые характеристики и вместо этого становится уплощенным с многослойными характеристиками. Профилирование экспрессии генов подтверждает, что транскриптом кишечника с дефицитом CDX2 больше напоминает профиль пищевода/переднего желудка, а не профиль кишечника (Gao et al., 2009). Точно так же у Cdx2+/- животных могут развиваться кишечные полипы, которые содержат многослойный плоский эпителий типа передней кишки (Chawengsaksophak et al., 19).97).
Роль CDX2 на более поздних стадиях развития кишечного эпителия обычно сохраняется, поскольку потеря CDX2 в кишечнике мышей, начиная с ~E13. 5, также приводит эпителий кпереди, хотя и частично. При элиминации на этой более поздней стадии CDX2-дефицитный кишечный эпителий экспрессирует SOX2, но не p63, и частично трансформируется в сторону идентичности типа железистого желудка, а не приобретает полностью стратифицированную идентичность пищевода/переднего желудка (Grainger et al., 2010). Точно так же индуцируемый нокаут Cdx2 эпителия кишечника взрослых индуцирует экспрессию маркеров эпителиальных клеток желудка и снижает экспрессию маркеров эпителиальных клеток кишечника (Stringer et al., 2012). Возможно, неспособность индуцировать р63 лежит в основе различий между степенью передней трансформации, наблюдаемой у ранних и поздних мутантов CDX2. CDX2 также абсолютно необходим для развития толстой кишки, поскольку условная делеция CDX2 в средней и задней кишке во время раннего эмбрионального развития полностью отменяет развитие толстой кишки (Gao et al., 2009).).
В совокупности функцию CDX2 можно сопоставить с функцией p63 и SOX2 при регионаризации желудочно-кишечного эпителия (). CDX2, высоко экспрессируемый в средней и задней кишке, необходим для прямого развития столбчатого кишечного эпителия, тогда как SOX2 и p63, оба высоко экспрессируются в передней кишке, необходимы для прямого развития многослойного эпителия пищевода/переднего желудка. Неудивительно наблюдение, что CDX2 присутствует при столбчатой метаплазии кишечного типа в пищеводе и желудке (Bai et al., 2002; Phillips et al., 2003). Более того, недавняя работа лаборатории Que предполагает механистическую связь между CDX2 и BE, поскольку экспрессия CDX2 сама по себе индуцирует кишечную метаплазию типа пищевода Барретта в SCJ in vivo у мышей (Jiang et al., 2017). Поразительное различие между эффектами ранней потери CDX2 в развивающейся тонкой кишке по сравнению с толстой кишкой может быть объяснено различными воздействиями на экспрессию ключевых нижестоящих эффекторов. Например, факторы передней кишки SOX2, PAX9, и WNT10A сильно индуцируются в дистальных отделах средней/задней кишки мутантов CDX2, а столбчатый фактор Indian Hedgehog (обсуждается ниже) только сильно снижен в дистальных отделах средней/задней кишки мутантов (Gao et al. , 2009).
iv. Транскрипционные факторы семейства Hepatic Nuclear Factor 4 в развитии столбчатого эпителия.
Семейство транскрипционных факторов Hepatic Nuclear Factor 4 (HNF4), которое включает HNF4 альфа (HNF4A) и HNF4 гамма (HNF4G), также участвует в контроле регионаризации эпителия ЖКТ (). HNF4A экспрессируется в развивающемся и зрелом эпителии желудка, тонкой и толстой кишки, тогда как HNF4G экспрессируется только в развивающейся и зрелой толстой кишке (Drewes et al., 19).96; Дункан и др., 1994; Ли и др., 2009 г.; Таравирас и др., 1994; Таравирас и др., 2000). Роль HNF4A в развитии эпителия толстой кишки исследовали с помощью условного нокаута (Garrison et al., 2006). Поскольку эта модель также удаляет HNF4A в печени, вызывая летальный фенотип (Battle et al., 2006; Parviz et al., 2003), фенотип толстой кишки может быть изучен только у эмбрионов. Элиминация HNF4A внутри эмбрионального эпителия толстой кишки серьезно нарушает архитектуру толстой кишки с дефектами как эпителиального морфогенеза, так и цитодифференцировки эпителиальных клеток (Garrison et al. , 2006). Несколько групп исследовали функцию HNF4A в формировании паттерна эпителия тонкой кишки на более поздних стадиях развития и у взрослых мышей, но ни одна из них не сообщила о каком-либо существенном влиянии на развитие эпителия тонкой кишки (Ahn et al., 2008; Babeu et al., 2009).; Дарсиньи и др., 2009). Возможно, что делеция HNF4A ранее в ходе развития тонкой кишки высветит новую роль HNF4A в этом процессе. Также возможно, что HNF4G, отсутствующий в эпителии толстой кишки, но присутствующий в эпителии тонкой кишки, компенсирует функцию HNF4A у мутантов. О роли HNF4A в развитии эпителия желудка не сообщалось. Однако группа Mills показала, что HNF4A необходим для цитодифференцировки и гомеостаза желудочного эпителия взрослых мышей (Moore et al., 2016). Хотя развитие желудочно-кишечного эпителия еще предстоит полностью изучить у мышей с нокаутом HNF4G, HNF4G участвует в регуляции развития границы желудка и двенадцатиперстной кишки. Экспрессия HNF4G резко повышается в эпителии двенадцатиперстной кишки по сравнению с таковой в желудке во время сегрегации этих доменов энтодермы передней кишки, что указывает на его роль в этом процессе (Li et al. , 2009).). Более того, было показано, что HNF4G активируется при метапластических поражениях желудка, что дополнительно подтверждает роль высоких уровней HNF4G в различении желудочного и кишечного доменов (Sousa et al., 2016).
v. GATA4 и GATA6 при регионализации цилиндрического эпителия желудка и кишечника.
Из шести известных транскрипционных факторов семейства GATA GATA4, GATA5 и GATA6 экспрессируются в развивающейся энтодерме, которая дает начало эпителию желудочно-кишечного тракта. Было показано, что GATA4 и GATA6, но не GATA5, являются важными регуляторами развития эпителия ЖКТ (Aronson et al., 2014; Battle et al., 2008; Beuling et al., 2008; Beuling et al., 2010; Beuling et al. al., 2011; Beuling et al., 2012; Bosse et al., 2006; Kohlnhofer et al., 2016; Thompson et al., 2017; Walker et al., 2014a; Walker et al., 2014b). В то время как GATA6 экспрессируется в эпителии железистого заднего отдела желудка, всего тонкого кишечника и толстой кишки, экспрессия GATA4 прекращается на границе тощей/подвздошной кишки в пределах эпителия тонкой кишки () (Battle et al. , 2008; Beuling et al., 2012; Bosse et al., 2006; Fang et al., 2006; Jacobsen et al., 2002). Исследования генетического анализа мозаики с химерными эмбрионами, содержащими Gata4 нулевые клетки указывают на важную роль GATA4 в обеспечении правильного развития столбчатого железистого эпителия в заднем желудке мышей (Jacobsen et al., 2002). В этой модели Gata4 нулевые клетки редко вносят вклад в железистый эпителий заднего желудка, указывая на то, что GATA4 является критическим регулятором общего развития заднего желудка. Несколько Gata4 нулевых областей, появляющихся внутри химерных задних желудков, выступают вперед, приобретая эпителиальную морфологию многослойного плоскоклеточного типа. Кроме того, 9В нулевых клетках 0128 Gata4 отсутствует экспрессия маркеров дифференцировки желудка. Было показано, что GATA4 взаимодействует с транскрипционным кофактором Friend-of-GATA (FOG1), чтобы направлять экспрессию генов в эпителии заднего отдела желудка (Jacobsen et al. , 2005). Условные нокауты для изучения роли GATA4 в регионализации желудка или исследования для изучения роли GATA6 в определении эпителия желудка еще не сообщались. Однако сообщалось об экспрессии GATA4 и GATA6 при пищеводе Барретта и аденокарциноме пищевода, что дает основание для дальнейших исследований этих факторов в регуляции регионализации плоскостолбчатого соединения (Duggan et al., 2016; Haveri et al., 2008; Miller et al. др., 2003).
Внутри доменов кишечника и толстой кишки факторы GATA в первую очередь контролируют регионализацию эпителиальной цитодифференцировки, а не структуру эпителия, хотя ранняя делеция GATA4 в средней кишке задерживает начало морфогенеза ворсинок (Kohlnhofer et al., 2016). Эти фенотипы цитодифференцировки обсуждаются ниже в разделе 3.
vi. Передача сигналов Hedgehog в эпителиальной регионализации ЖКТ.
Белки Hedgehog (HH), секретируемые эпителиальными клетками, действуют на нижележащие мезенхимальные клетки, которые, в свою очередь, посылают обратно эпителию сигналы для направления его развития (van den Brink, 2007). Два белка HH, Sonic Hedgehog (SHH) и Indian Hedgehog (IHH), динамически экспрессируются в развивающейся энтодерме и ее производных эпителиях (Feng et al., 2012; Khurana and Mills, 2010; Ramalho-Santos et al., 2000; ван ден Бринк, 2007). В начале развития SHH и IHH широко экспрессируются в энтодерме передней, средней и задней кишки. Позднее SHH преимущественно экспрессируется в доменах пищевода и преджелудка, тогда как IHH в основном экспрессируется в доменах заднего желудка и кишечника (10). К E18.5, после того как эпителиальные домены ЖКТ претерпели значительное морфогенетическое ремоделирование, экспрессия SHH снижается в массе многослойного плоского эпителия пищевода, ограничиваясь дистальным эпителием пищевода, но увеличивается в эпителии заднего отдела желудка и кишечника. Этот динамический временной паттерн экспрессии белков HH имеет важные разветвления для регионализации эпителия GI (16).
Открыть в отдельном окне
Критическая граница передачи сигналов hedgehog необходима для регионализации эпителия желудка мыши в области плоскостолбчатого соединения. На ранних стадиях развития Sonic Hedgehog (SHH) и Indian Hedgehog (IHH) широко экспрессируются во всех трех областях энтодермы, передней кишке (FG), средней кишке (MG) и задней кишке (HG). По мере прогрессирования развития SHH ограничивается доменами, дающими начало многослойному плоскому эпителию (пищевод, E, и преджелудок, FS), а IHH ограничивается доменами, дающими начало столбчатому эпителию (задний желудок, HS, двенадцатиперстная кишка, D, тощая кишка, J, подвздошной кишки, I и толстой кишки, C). Граница экспрессии SHH/IHH в развивающемся желудке является критической для правильного развития SCJ. При нарушении границы многослойного плоского и цилиндрического эпителия смещаются. Потеря SHH в области преджелудка с сопутствующей индукцией IHH приводит к задним изменениям эпителия преджелудка. Наоборот, потеря IHH в заднем отделе желудка с сопутствующей индукцией SHH приводит к переднему краю заднего отдела желудка. В таблице суммированы фенотипы, наблюдаемые у мутантов мышей с приобретением и потерей функции HH в желудочно-кишечном тракте.
Исследования усиления и потери функции SHH в пищеводе выявили существенную роль этого сигнального пути в регионаризации эпителия пищевода. Полное отсутствие SHH в ранней развивающейся энтодерме передней кишки блокирует разделение трахеи и пищевода, вызывая трахеопищеводные свищи (Litingtung et al., 1998). Более недавнее исследование любых оставшихся клеток пищевода у мутантов SHH показало, что устранение SHH в домене пищевода приводит к преждевременной экспрессии маркеров стратифицированных эпителиальных клеток (Wang et al., 2014). Эти данные свидетельствуют о том, что хотя подавление SHH вредно на самых ранних стадиях развития пищевода, подавление SHH на более поздних стадиях является ключом к контролю начала программы дифференцировки многослойных плоскоклеточных эпителиальных клеток в пищеводе. Дальнейшим подчеркиванием необходимости подавления SHH в домене пищевода для обеспечения развития стратифицированных эпителиальных клеток является обнаружение того, что реактивация SHH в эпителии пищевода вызывает развитие столбчатых эпителиальных признаков, одним из которых является экспрессия регулируемой SHH транскрипции. фактор FOXA2 (Wang et al., 2014). И SHH, и FOXA2 повторно активируются при столбчатой метаплазии пищевода Барретта (Wang et al., 2014), что указывает на то, что передача сигналов SHH вредна для гомеостаза многослойного плоского эпителия пищевода.
Возможно, неудивительно, что передача сигналов HH также является важным игроком в регионаризации эпителия желудка (16). Желудки с отсутствием SHH обнаруживают сильно расширенный железистый столбчатый домен желудка за счет многослойного плоского домена преджелудка (Ramalho-Santos et al., 2000). Более того, железистый желудок мутантов SHH также апостериоризован, экспрессируя некоторые кишечные маркеры. Сходным образом потеря HOXA5 из нижележащей желудочной мезенхимы нарушает эпителиальную передачу сигналов HH в развивающемся желудке. Экспрессия IHH расширяется в домен преджелудка за счет экспрессии SHH у мутантов HOXA5, и мутантные желудки апостериоризуются, экспрессируя кишечный маркер щелочную фосфатазу даже в презумптивном домене преджелудка (14) (Aubin et al. , 2002). С другой стороны, индуцированная экспрессия SHH в заднем желудке вследствие 9Нокаут 0128 Hnf1b , нокаут Fgf10 , нокаут Fgfr2 или экспрессия мутантных рецепторов активина нарушает домен IHH в заднем желудке и перемещает эпителий вперед () (Haumaitre et al., 2005; Kim et al., 2000; Спенсер-Ден и др., 2006). В этих случаях мутантный эпителий утолщается и происходит потеря типов железистых клеток. Взятые вместе, баланс между экспрессией SHH и IHH необходим для формирования паттерна многослойного плоского и столбчатого эпителия желудка в желудке (4).
Передача сигналов HH также регулирует критические аспекты эпителиального морфогенеза в тонком кишечнике (Madison et al., 2005; Walton et al., 2012; Walton et al., 2016a). Когда передача сигналов Hedgehog блокируется экспрессией белка-ингибитора pan-Hedgehog Hhip, эпителий остается незрелым с персистирующей псевдостратифицированной архитектурой (Madison et al., 2005). Более того, сигналы HH контролируют кластеризацию лежащих в основе PDGFRA+ мезенхимальных клеток, что абсолютно необходимо для разрастания ворсинок, и поэтому морфогенез ворсинок является аномальным у мутантов HH (Walton et al. , 2012). Наконец, передача сигналов HH регулирует локализацию ниши пролиферативных клеток в кишечных криптах. При блокировке пролиферативные клетки мигрируют вверх из крипт, образуя эктопические крипты, которые аномально разветвляются на ворсинки (Madison et al., 2005).
vii. Передача сигналов WNT в регионализации эпителия ЖКТ.
Во время развития передней кишки передача сигналов WNT должна быть сведена к минимуму для правильной регионализации. Если ранняя энтодерма передней кишки мыши (E7.5-8.5) подвергается воздействию передачи сигналов WNT посредством экспрессии конститутивно активного B-катенина, энтодерма передней кишки начинает аберрантно экспрессировать главный регулятор кишечника CDX2, как правило, за счет регулятора передней кишки SOX2, хотя и редко. Присутствуют коэкспрессирующие клетки CDX2, SOX2 (Sherwood et al., 2011). Более того, экспрессия CDX2 в этих клетках достаточна, чтобы индуцировать программу экспрессии генов, подобную кишечной, что приводит к заднему развитию передней кишки.
Гомеодоменовый фактор транскрипции BarH-подобный гомеобокс 1 (BARX1), экспрессируемый в мезенхиме пищевода, преджелудка и заднего отдела желудка, играет центральную роль в ингибировании передачи сигналов WNT в заднем домене передней кишки (). Потеря мезенхимального BARX1 снижает экспрессию ингибиторов передачи сигналов WNT, секретируемых белками 1 и 2, связанными с frizzled (SFRP1/2) (Kim et al., 2005; Kim et al., 2007; Woo et al., 2011). Аномальная активация передачи сигналов WNT в энтодерме передней кишки BARX1-дефицитных желудков оказывает влияние на домены органов пищевода, преджелудка и заднего отдела желудка (Kim et al., 2005; Kim et al., 2007). Вся передняя кишка с мутациями BARX1 является задней, с передними (пищевод/передний желудок) областями, содержащими эпителий смешанного «чешуйчато-железистого» типа с пониженными уровнями SOX2. Возможно, наиболее поразительным является эпителий, подобный кишечным ворсинкам, присутствующий в задних желудках мутантов BARX1. В этой области аберрантная передача сигналов WNT индуцирует экспрессию CDX2, чтобы сместить глобальный профиль экспрессии генов в развивающемся желудке в сторону более задней программы средней кишки/кишечной энтодермы. Для мезенхимальной экспрессии родственного гомеобоксного фактора транскрипции, BAPX1, в дистальном отделе заднего отдела желудка требуется BARX1, а экспрессия дистальных маркеров желудка и сужение привратника отсутствуют в обоих случаях.0128 Barx1 и Bapx1 нулевые мыши (Verzi et al., 2009).
Открыть в отдельном окне
Передача сигналов WNT должна подавляться в передней кишке и индуцироваться в средней и задней кишке для правильной регионализации эпителия желудочно-кишечного тракта. В развивающемся желудке передача сигналов WNT должна быть ингибирована для правильного развития желудочного эпителия. Отсутствие WNT имеет решающее значение для установления экспрессии SOX2 в этом домене. Мезенхимальный фактор транскрипции BARX1 необходим для ингибирования передачи сигналов WNT в домене передней кишки. BARX1 индуцирует экспрессию ингибиторов WNT, известных как SFRP (секретируемые белки, родственные frizzled). В этой среде с низким WNT происходит правильное формирование многослойного плоского эпителия пищевода, преджелудка и железистого столбчатого эпителия заднего желудка. В отсутствие BARX1 активируется WNT. Эпителий пищевода и преджелудка приобретает «чешуйчато-железистую» гибридную структуру, тогда как эпителий заднего отдела желудка становится интестинизированным. BAPX1, др. желудочный мезенхимальный фактор, также способствует дифференцировке желудочного и кишечного доменов. В областях задней энтодермы разрешена передача сигналов WNT и индуцируется CDX2. В толстой кишке потеря эффекторов WNT TCF1 и TCF4 блокирует развитие задней части кишечника. Кроме того, для образования простого столбчатого эпителия толстой кишки необходимы перекрестные помехи между HH и WNT. При отсутствии ИГГ WNT нарушается, а эпителий антериоризуется. В таблице суммированы фенотипы в экспериментах с потерей и усилением функции с участием активаторов и репрессоров WNT, а также белков, участвующих в перекрестных разговорах с путем WNT, во время развития желудочно-кишечного тракта и морфогенеза эпителия.
Передача сигналов WNT также играет решающую роль в развитии толстой кишки. Экспрессия ингибитора WNT Dickkopt-1 (DKK-1) приводит к сильно измененной морфологии эпителия толстой кишки, включая потерю архитектуры крипт (Kuhnert et al., 2004). Кроме того, эмбрионы, лишенные эффекторов WNT TCF1 и TCF4, неспособны формировать заднюю кишку (Gregorieff et al., 2004). Кроме того, существует взаимодействие между сигнализацией HH и WNT, при этом сигнализация HH ограничивает передачу сигналов WNT базой шифрования. Соответственно, IHH-дефицитная эмбриональная толстая кишка лишена крипт и имеет многослойный эпителий, а не простой столбчатый эпителий (van den Brink et al., 2004).
viii. Костные морфогенетические белки (BMP) в регионализации эпителия ЖКТ.
Правильная временная модуляция передачи сигналов BMP абсолютно необходима во время развития пищевода и преджелудка (Que et al., 2006; Rodriguez et al., 2010). В течение раннего периода эпителиальной стратификации передача сигналов BMP должна быть ингибирована. Фактически, если передача сигналов BMP присутствует, она репрессирует экспрессию SOX2 в передней кишке, чтобы нарушить развитие эпителия пищевода (Domyan et al. , 2011). Чтобы защитить эпителий пищевода от мезенхимальных сигналов BMP, эпителий секретирует антагонисты BMP для блокирования передачи сигналов BMP (Que et al., 2006; Rodriguez et al., 2010). Экспрессия конститутивно активного рецептора BMP в развивающемся пищеводе и преджелудке блокирует расслоение эпителия (Rodriguez et al., 2010). Позже требуется активная передача сигналов BMP, поскольку эпителий пищевода, лишенный рецепторов BMP, не может дифференцироваться (Rodriguez et al., 2010). Интересно, что стромальная активация BMPs в пищеводе взрослых была связана с столбчатой метаплазией в пищеводе Барретта (Castillo et al., 2012).
Открыть в отдельном окне
Временная модуляция передачи сигналов BMP необходима для развития множественных эпителиев ЖКТ. (A) В начале развития передней кишки передача сигналов BMP должна быть ингибирована, чтобы индуцировать стратификацию эпителия, тогда как позже, в развитии пищевода и преджелудка, передача сигналов BMP должна быть активной, чтобы индуцировать дифференцировку супрабазальных клеток в многослойном плоском эпителии этих органов. (B) Передача сигналов BMP контролирует морфогенез ворсинок в тонкой кишке посредством структурирования кластеров мезенхимальных PDGFRA+ клеток. Мезенхимальные клетки под развивающимся кишечным эпителием (i) реагируют на передачу сигналов BMP, кластер (ii) и происходит разрастание ворсинок (iii). Скопления мезенхимы остаются на кончике ворсинки по мере того, как ворсинки продолжают развиваться (iii). (C) Дифференцировка индуцированных человеком плюрипотентных стволовых (iPS) клеток в органоиды тонкого кишечника управляется передачей сигналов WNT и FGF, в то время как получение колоноидов из iPS-клеток требует передачи сигналов WNT, FGF и BMP. В таблице суммированы фенотипы, обусловленные потерей и приобретением функций, исследования с участием ключевых участников передачи сигналов BMP во время развития органов желудочно-кишечного тракта.
В тонком кишечнике передача сигналов BMP играет ключевую роль в морфогенезе ворсинок. Однако передача сигналов BMP в первую очередь сосредоточена внутри мезенхимальных кластеров, а не в эпителии (Walton et al. , 2016b). В то время как делеция рецепторов BMP в эпителии не может нарушить формирование ворсинок, модуляция передачи сигналов BMP в мезенхиме — повышение или понижение мезенхимального BMP — нарушает морфогенез ворсин путем изменения структуры мезенхимальных PDGFRA+ клеточных кластеров (4). Передача сигналов BMP также необходима для развития толстой кишки из энтодермы задней кишки. Недавние исследования с использованием плюрипотентных стволовых клеток человека показали, что в присутствии передачи сигналов WNT, FGF и BMP развиваются органоиды толстой кишки человека; однако в отсутствие BMP по умолчанию используются органоиды тонкой кишки (Múnera et al., 2017).
Первая часть этого обзора была сосредоточена на том, как факторы транскрипции и пути передачи сигналов вносят вклад в региональное формирование паттерна трехмерной эпителиальной структуры. Теперь мы рассмотрим другой ключевой аспект регионализации эпителия ЖКТ, а именно механизмы, с помощью которых развиваются существенные органоспецифические типы эпителиальных клеток. Мы сосредоточимся на регионализации типов эпителиальных клеток в железистом заднем желудке и тонкой кишке (2).
Открыть в отдельном окне
Ключевые факторы транскрипции и сигнальные молекулы, контролирующие цитодифференцировку в эпителии желудка и тонкого кишечника.
A) На иллюстрации сравнивается клеточная архитектура проксимальных (фундальных) и дистальных (антральных) желез заднего отдела желудка и показаны ключевые транскрипционные факторы и сигнальные молекулы, участвующие в регуляции цитодифференцировки в этих областях. Передача сигналов WNT необходима для определения желез проксимального (фундального) типа и отсутствует в дистальной (антральной) области. И наоборот, дистальные (антральные) железы экспрессируют фактор транскрипции PDX1, тогда как проксимальные (фундальные) железы не содержат PDX1. Обратите внимание, что париетальные клетки, главные клетки и клетки слизи шеи обогащены проксимальными (фундальными) железами по сравнению с дистальными (антральными) железами. B) Иллюстрация изображает клеточную архитектуру ворсинок и крипт тонкой кишки и показывает ключевые факторы транскрипции и сигнальные молекулы, участвующие в регуляции кишечной цитодифференцировки. Факторы обозначаются как D (двенадцатиперстная кишка), J (тощая кишка) и/или I (подвздошная кишка), чтобы описать характер их экспрессии. Обратите внимание, что энтероциты регионализованы по дифференциальной экспрессии PDX1 (D) и GATA4 (D, J), а энтероэндокринные клетки регионализированы по дифференциальной экспрессии PAX4 (D, J), PAX6 (D, J) и PDX1 (D).
A. Желудочная цитодифференцировка.
Определенные типы эпителиальных клеток желудка синтезируют кислоту, пищеварительные ферменты и гормоны, которые обеспечивают химическое и механическое переваривание пищи и регулируют моторику желудка () (Choi et al., 2014; Kim and Shivdasani, 2016; McCracken and Wells, 2017; Willet и Миллс, 2016). Два типа клеток секретируют слизь: клетки слизи шеи и клетки поверхностной слизи (ямки/фовеолярные клетки). Главные клетки секретируют пищеварительные ферменты, в том числе пепсиноген. Париетальные клетки выделяют кислоту для контроля рН желудка. Энтероэндокринные клетки секретируют гормоны в нижележащую собственную пластинку пластинки, такие как гастрин, для стимуляции секреции кислоты. Наконец, клетки желудочного пучка, являясь типом люминальных химиочувствительных клеток, проецируют апикальные отростки в просвет, а также продуцируют секретируемые молекулы, в том числе ацетилхолин, для стимуляции секреции эпителиальных клеток, и цитокины, чтобы опосредовать воспалительные реакции (Gerbe and Jay, 2016; Hayakawa et al. , 2017; Каджи и Кауниц, 2017). Распределение определенных типов эпителиальных клеток в данной железе желудка варьируется вдоль проксимальной и дистальной оси железистой задней части желудка () (Choi et al., 2014; Kim and Shivdasani, 2016; McCracken and Wells, 2017; Willet and Mills, 2016). У грызунов все типы дифференцированных клеток присутствуют в железах проксимального (также известного как фундальный) заднего отдела желудка, тогда как главные клетки и париетальные клетки отсутствуют в дистальном (также известном как антральный) заднем отделе желудка. Гастрин-продуцирующие энтероэндокринные клетки (G-клетки) находятся исключительно в дистальных (антральных) железах заднего желудка у мышей и людей. Дистальные (антральные) железы также содержат клетки базальных желез, которые демонстрируют черты как шейной слизи, так и главных клеток (Willet and Mills, 2016). Хотя есть примерно 90% снижение париетальных клеток вдоль переходной зоны между проксимальными (фундальными) желудочными железами и дистальными (антральными) желудочными железами в желудке человека, железы, содержащие как G-клетки, экспрессирующие гастрин, так и париетальные клетки, продуцирующие кислоту, могут существовать в самых дистальных отделах. желудка (Choi et al., 2014).
Хотя механизмы, контролирующие цитодифференцировку эпителия в желудке, менее изучены по сравнению с кишечником, несколько транскрипционных факторов и сигнальных путей стали играть ключевую роль. Передача сигналов WNT играет существенную роль в регионализации идентичности типов эпителиальных клеток в пределах проксимального (фундального) и дистального (антрального) доменов железистого заднего желудка. Одним из ключевых маркеров, отличающих эти домены, является транскрипционный фактор панкреатического и дуоденального гомеобокса 1 (PDX1), который присутствует в дистальных (антральных) железах, но отсутствует в проксимальных (фундальных) железах. Когда передача сигналов WNT блокируется на ранней стадии в развивающемся проксимальном эпителии заднего отдела желудка путем элиминации B-катенина с использованием Shh-Cre, экспрессия маркера проксимальной железы заменяется экспрессией маркера дистальной железы на E10.5, включая индукцию PDX1 (McCracken et al., 2017). Это указывает на то, что передача сигналов WNT необходима для спецификации проксимальных и дистальных типов желудочных желез во время развития. По аналогии с желудком мыши, передача сигналов WNT также необходима для стимуляции развития проксимальных (фундальных) железистых отделов желудка, что продемонстрировано с использованием органоидов желудка, полученных из плюрипотентных клеток человека (McCracken et al., 2017). Когда WNT добавляют к культурам во время фазы спецификации желудка, появляются органоиды проксимального (фундального) типа. Исключение WNT позволяет формировать органоиды дистального (антрального) типа. В совокупности эти данные указывают на то, что регуляция передачи сигналов WNT важна для правильной регионализации желудочных желез у мышей и людей.
Для созревания главных клеток в проксимальном (фундальном) отделе заднего желудка требуется фактор транскрипции, связывающий X-box белок 1 (XBP1), и его мишень, мышечная экспрессия кишечника и желудка 1 (MIST1) (Huh et al., 2010; Ramsey et al. , 2007; Тиан и др., 2010). Совсем недавно MIST1 был идентифицирован как фактор транскрипции, который широко контролирует установление и поддержание генерализованной секреторной клеточной архитектуры в различных типах клеток, включая главную клетку желудка (Lo et al., 2017). У мышей с нулевым эстроген-родственным рецептором гамма (ESRRG) снижено количество париетальных клеток в проксимальных (фундальных) железах, что свидетельствует о регулируемой ESRRG продукции этих клеток (Alaynick et al., 2010). Дифференцировка пит-клеток нуждается в белке FOXQ1 с вильчатой головкой (Verzi et al. , 2008). GATA4 может также играть роль в развитии типов клеток желудка, включая париетальные клетки, главные клетки и клетки, продуцирующие слизь шеи (Jacobsen et al., 2002).
Типы энтероэндокринных клеток варьируют от проксимального до дистального отдела заднего желудка, и специфические факторы транскрипции регулируют дифференцировку типов, специфичных для региона. Например, нейрогенин 3 (NGN3) и ARX необходимы для дифференцировки различных подмножеств энтероэндокринных клеток желудка, обнаруживаемых либо в проксимальной (фундальной), либо в дистальной (антральной) областях (Du et al., 2012; Jenny et al., 2002; Кокубу и др., 2008; Ли и др., 2002). PDX1, NKX6.3 и PAX6 необходимы специфически в дистальном (антральном) заднем отделе желудка для дифференцировки G-клеток, продуцирующих гастрин, тогда как PAX4 необходим для энтероэндокринных подтипов, продуцирующих серотонин и соматостатин в этой области (Choi et al., 2008; Ларссон и др., 1996; Ларссон и др., 1998). Клетки пучка, хотя и присутствуют по всему заднему желудку, обогащены в области плоскостолбчатого соединения и нуждаются в Pou2f3 (Gerbe et al. , 2016). SPDEF необходим для дифференцировки клеток слизи в дистальном (антральном) заднем отделе желудка (Horst et al., 2010).
Наконец, потеря передачи сигналов BMP желудка нарушает развитие париетальных клеток в проксимальных (фундальных) железах (Shinohara et al., 2010). Эпителиальная гиперплазия и развитие гибридных клеток, продуцирующих как муцины, так и зимогены, также происходят в отсутствие надлежащей передачи сигналов BMP. В целом, передача сигналов BMP, по-видимому, играет жизненно важную роль в поддержании эпителия желудка, поскольку потеря передачи сигналов BMP приводит к метаплазии и дисплазии (Todisco, 2017).
B. Кишечная цитодифференцировка.
Подобно желудочному эпителию, определенные типы эпителиальных клеток кишечника работают согласованно, чтобы выполнять функции тонкой кишки. Решения о судьбах клеток, которые определяют спецификацию секреторных и абсорбирующих эпителиальных клеток кишечника, были тщательно изучены и подробно рассмотрены другими (Bjerknes and Cheng, 2010; Cheng and Leblond, 1974; Chin et al. , 2017; De Mey and Freund, 2013; Noah et al., 2011; Qi and Chen, 2015; Simons and Clevers, 2011; Spence et al., 2011). Линия секреторных клеток состоит из четырех подтипов: бокаловидных, энтероэндокринных, пучковых и клеток Панета. Бокаловидные клетки составляют примерно 10% клеток кишечного эпителия. Эти клетки продуцируют и секретируют муцины, которые покрывают апикальную поверхность эпителия, обеспечивая защитный барьер от люминальных патогенов (Birchenough et al., 2015; McCauley and Guasch, 2015). Энтероэндокринные клетки, составляющие только 1% кишечного эпителия, секретируют гормоны в собственную пластинку пластинки для регуляции процессов, включая рост и восстановление клеток, гликемию, экзокринную секрецию поджелудочной железы и подвижность стенок кишечника (Jenny et al., 2002). Клетки пучка, составляющие редко 0,4% эпителия, секретируют опиоиды для контроля боли, опорожнения желудка, кишечной секреции и перистальтики кишечника (Gerbe et al., 2012; Gerbe and Jay, 2016; Holzer, 2009). ; Каджи и Кауниц, 2017). Клетки Панета, секретирующие антимикробные пептиды в просвет, являются единственным типом зрелых клеток, которые мигрируют вниз в крипту, а не вверх вдоль ворсинки (Andreu et al., 2008; Ayabe et al., 2000). Наконец, наиболее многочисленны клетки абсорбционного ряда, энтероциты. Энтероциты имеют микроворсинки на своей апикальной поверхности, что значительно увеличивает площадь поверхности просвета, что способствует максимальной эффективности поглощения питательных веществ.
Из этих типов клеток как энтероциты, так и энтероэндокринные клетки специализированы по длине эпителия тонкой кишки, выполняя специфические функции в различных отделах кишечника (). Механизм(ы) передачи сигналов и факторы транскрипции, необходимые для регионализации энтероцита, придающего ему различные функции, изучены недостаточно. Факторы, по-разному экспрессируемые в энтероцитах в этих областях, вероятно, играют жизненно важную роль в определении и поддержании региональной идентичности энтероцитов. PDX является одним из таких факторов. Экспрессия PDX1 охватывает границу SOX2/CDX2, простирающуюся от дистального (антрального) SOX2+ заднего отдела желудка до CDX2+ двенадцатиперстной кишки, что делает PDX1 специфическим для двенадцатиперстной кишки фактором транскрипции в тонкой кишке (19) (Guz et al., 19).95). У мышей, лишенных PDX1, нарушена абсорбция железа, специфическая функция двенадцатиперстной кишки, а также нарушена абсорбция липидов и клиренс липидов (Chen et al., 2009). В соответствии с этим открытием сверхэкспрессия PDX1 в клеточной линии Caco-2, полученной из карциномы толстой кишки, вызывает изменения в экспрессии генов, связанных с метаболизмом питательных веществ и липидов (Chen et al., 2012a). Гены, непосредственно регулируемые PDX1, включают маркеры проксимальных энтероэндокринных клеток, ингибирующий полипептид желудка (Gip) и соматостатин (Sst), а также экспрессируемый в двенадцатиперстной кишке ген Ada (Chen et al., 2009).; Дусинг и др., 2001; Джепил и др., 2005). Также было показано, что PDX1 подавляет транскрипцию сахаразы, изомальтазы, лактазы и белка 1, связывающего жирные кислоты, каждый из которых кодирует белки, в первую очередь связанные с функцией тощей кишки (Chen et al., 2012b; Heller et al., 1998; Wang et al., 2004). . Эти данные указывают на то, что PDX1 является ключевым регулятором идентичности энтероцитов двенадцатиперстной кишки.
Другим дифференциально экспрессируемым транскрипционным фактором является GATA4, который присутствует в эпителии двенадцатиперстной и тощей кишки, но отсутствует в эпителии подвздошной кишки (Battle et al., 2008; Bosse et al., 2006; Fang et al., 2006). . Ранний (E11.5) условный нокаут Gata4 в кишечнике приводит к дефекту пролиферации эпителиальных клеток посредством регуляции транскрипции белков клеточного цикла и рецептора 5 класса Frizzled ( Fzd5 ) (Kohlnhofer et al., 2016). Кроме того, задерживается морфогенез ворсинок и изменяются популяции эпителиальных клеток. Важно отметить, что транскрипты, обогащенные тощей кишкой, уменьшаются, а транскрипты, обогащенные подвздошной кишкой, увеличиваются. Сдвиг от тощей кишки к подвздошной идентичности также происходит, когда GATA4 удаляется в кишечном эпителии на более поздних стадиях развития (E16.5) с использованием Villin-Cre (Walker et al., 2014b). Морфологически тощая кишка кишечника с нокаутом E16.5 GATA4-Villin-Cre преимущественно нормальная, но с измененной экспрессией генов в популяциях энтероцитов и энтероэндокринных клеток и небольшим увеличением числа бокаловидных клеток. Изменения в экспрессии генов тощекишечных энтероцитов, наблюдаемые у эмбриональных мутантов GATA4, сохраняются во взрослом возрасте, при этом GATA4-дефицитная тощая кишка теряет экспрессию 53% набора генов, специфичных для тощей кишки, и приобретает экспрессию 47% набора генов, специфичных для подвздошной кишки (Battle et al. , 2008). Важно, что репрессированный набор специфичных для тощей кишки включает гены, кодирующие белки, необходимые для транспорта липидов и холестерина. Подвздошно-специфический набор включает гены, кодирующие белки, участвующие в абсорбции желчных кислот. Следовательно, эти мыши поглощают меньше жира и холестерина, чем мыши дикого типа. Индуцируемый нокаут GATA4 из зрелого эпителия приводит к сходному эффекту на экспрессию генов (Bosse et al., 2006). Совсем недавно было показано, что GATA4 необходим и достаточен для поддержания тощей кишки и подавления подвздошной идентичности в эпителии тонкой кишки (Thompson et al., 2017). Эктопическая экспрессия GATA4 в подвздошной кишке мыши приводит к потере программы экспрессии генов подвздошной кишки и усилению программы тощей кишки. Вместе эти данные указывают на то, что GATA4 необходим для установления и поддержания идентичности тощей и подвздошной энтероцитов. На сегодняшний день GATA4 является единственным известным фактором, играющим столь важную роль в регуляции границы тощей и подвздошной кишек.
Важно отметить, что GATA6 присутствует во всем эпителии кишечника и что присутствие GATA6 в тощей кишке с дефицитом GATA4 или в подвздошной кишке, экспрессирующей GATA4, не компенсирует потерю или усиление функции GATA4 в этих тканях (Walker et al. , 2014а; Уокер и др., 2014b). Более того, индуцируемая делеция GATA6 из эпителия кишечника взрослых не изменяет идентичности тощекишечных энтероцитов (Beuling et al., 2011). Вместе эти данные подчеркивают, что регионализация тощекишечных энтероцитов является функцией, уникальной для GATA4. Когда GATA6 удален из подвздошной кишки (где GATA4 отсутствует), однако, экспрессия генов подвздошных энтероцитов изменяется с экспрессией маркеров эпителиальных клеток толстой кишки Car1 и Car2, индуцированной (Beuling et al., 2011). Потеря GATA6 из эмбрионального эпителия подвздошной кишки также снижает экспрессию маркеров подвздошных энтероцитов и индуцирует экспрессию маркера эпителиальных клеток толстой кишки Car1 (Walker et al., 2014b). Одновременная индуцируемая делеция как GATA4, так и GATA6 из эпителия кишечника взрослых приводит к тому, что эпителиальные клетки двенадцатиперстной кишки и тощей кишки проявляют фенотип, сходный с GATA6-дефицитным подвздошным белком (Beuling et al., 2011). Сходный фенотип также встречается у эмбрионов, лишенных как GATA4, так и GATA6 в тощекишечном эпителии (Walker et al. , 2014a). Эти данные свидетельствуют о том, что в то время как GATA4 контролирует формирование паттерна тощекишечных и подвздошных энтероцитов, GATA6 уточняет идентичность подвздошных энтероцитов в кишечном эпителии путем репрессии экспрессии генов толстой кишки.
Отдельные подтипы энтероэндокринных клеток также по-разному распределяются по отделам тонкой кишки. Например, GIP-экспрессирующие и CCK-экспрессирующие энтероэндокринные клетки преобладают в проксимальном отделе кишечника, в то время как PYY-экспрессирующие энтероэндокринные клетки преобладают в дистальном отделе кишечника (Schonhoff et al., 2004). Было обнаружено, что несколько факторов транскрипции играют ключевую роль в развитии энтероэндокринных клеток, включая ATOh2, NGN3, NEUROD1, PDX1, PAX4 и PAX6 (Schonhoff et al., 2004; Sheaffer and Kaestner, 2012). В то время как ATOh2, NGN3 и NEUROD1, по-видимому, регулируют глобальное развитие энтероэндокринных клеток в кишечнике, PDX1, PAX4 и PAX6 контролируют энтероэндокринную дифференцировку в проксимальном отделе кишечника, при этом энтероэндокринные клетки практически исчезают из двенадцатиперстной и тощей кишки мутантов () (Larsson et al. ., 1998; Шонхофф и др., 2004; Шиффер и Кестнер, 2012). Различия в популяциях регионально обогащенных энтероэндокринных клеток также встречаются в тонком кишечнике условно нокаутных эмбрионов GATA4 и GATA6 (Battle et al., 2008; Walker et al., 2014a; Walker et al., 2014b). В то время как GATA6 присутствует в энтероэндокринных клетках и, следовательно, может осуществлять прямой контроль над дифференцировкой клеток, GATA4 отсутствует в этой клеточной популяции (Bosse et al., 2006; Walker et al., 2014b). Следовательно, GATA4, вероятно, косвенно регулирует популяцию энтероэндокринных клеток в кишечнике.
Хотя горстка сигнальных путей и транскрипционных факторов идентифицирована как ключевые регуляторы регионального паттернирования желудочно-кишечного тракта, для полноты картины необходима дополнительная работа. Ожидается, что полное понимание механизмов, с помощью которых эпителий ЖКТ приобретает и поддерживает уникальные паттерны экспрессии генов и архитектуру тканей, необходимые для региональных специфических функций, обеспечит существенное новое понимание заболеваний ЖКТ, включая метаплазии, рак и синдром короткой кишки. Например, знание нормальных механизмов развития и гомеостаза эпителия пищевода и желудка обеспечивает основу для разработки новых стратегий диагностики, лечения и ингибирования метаплазии, включая пищевод Барретта, метаплазию, экспрессирующую спазмолитические полипептиды, и кишечную метаплазию в желудке. Знание механизмов формирования паттерна, которые регулируют идентичность эпителиальных клеток двенадцатиперстной, тощей и подвздошной кишки в тонкой кишке, обеспечивает основу для будущей разработки лучших методов лечения синдрома короткой кишки, особенно тех, которые направлены на восстановление утраченных функций оставшейся кишечной ткани. Наконец, знание механизмов паттернирования эпителия ЖКТ было использовано для разработки новых и уточнения существующих in vitro модельных систем органов ЖКТ, в том числе полученных путем направленной дифференцировки плюрипотентных стволовых клеток или полученных непосредственно из образцов тканей человека. Эти модели имеют огромные перспективы для улучшения нашего понимания самого развития желудочно-кишечного тракта человека, для улучшения наших знаний об инфекционных заболеваниях, для улучшения фармакологических и токсикологических исследований, а также для инновационных подходов тканевой инженерии и персонализированной медицины к лечению желудочно-кишечных заболеваний.
Особенности
Регионализация эпителия желудочно-кишечного тракта необходима для эффективного усвоения питательных веществ.
Развитие уникальных органоспецифических эпителиальных структур и типов клеток необходимо для того, чтобы органы выполняли различные пищеварительные функции.
Пространственно-временная регуляция сигнальных путей и нижестоящих факторов транскрипции контролирует морфогенез и цитодифференцировку эпителия ЖКТ.
В этом обзоре обсуждаются фундаментальные механизмы, управляющие формированием паттерна желудочно-кишечного эпителия вдоль цефалокаудальной оси ЖКТ для обеспечения региональных специфических функций.
Финансирование: Эта работа была поддержана Национальными институтами здравоохранения, Национальным институтом диабета и болезней органов пищеварения и почек (номера грантов DK087873 и DK111822, MAB).
Отказ от ответственности издателя: Это PDF-файл неотредактированной рукописи, которая была принята к публикации. В качестве услуги нашим клиентам мы предоставляем эту раннюю версию рукописи. Рукопись будет подвергнута редактированию, набору текста и рецензированию полученного доказательства, прежде чем она будет опубликована в ее окончательной цитируемой форме. Обратите внимание, что в процессе производства могут быть обнаружены ошибки, которые могут повлиять на содержание, и все правовые оговорки, применимые к журналу, относятся к нему.
- Ан С.Х., Шах Ю.М., Иноуэ Дж., Моримура К., Ким И., Йим С., Ламберт Г., Куротани Р., Нагашима К., Гонсалес Ф.Дж., Иноуэ И., 2008. Ядерный фактор гепатоцитов 4альфа в эпителиальных клетках кишечника защищает от воспалительных заболеваний кишечника. Воспаление кишечника Dis 14, 908–920. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Алейник В. А., Уэй Дж. М., Уилсон С. А., Бенсон В. Г., Пей Л., Даунс М. , Ю Р., Йонкер Дж. В., Холт Дж. А., Раджпал Д. К., Ли Х., Стюарт Дж., Макферсон Р., Ремлингер К.С., Чанг С.И., Макдоннелл Д.П., Эванс Р.М., Биллин А.Н., 2010.
ERRgamma регулирует сердечный, желудочный и почечный гомеостаз калия. Мол Эндокринол
24, 299–309. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Andreu P, Peignon G, Slomianny C, Taketo MM, Colnot S, Robine S, Lamarque D, Laurent-Puig P, Perret C, Romagnolo B, 2008. Генетическое исследование роли передачи сигналов Wnt/бета-катенина в дифференцировке клеток Панета. Дев Биол 324, 288–296. [PubMed] [Google Scholar]
- Аронсон Б.Е., Рабелло Аронсон С., Берхаут Р.П., Чавуши С.Ф., Хе А., Пу В.Т., Верци М.П., Красински С.Д., 2014. GATA4 подавляет илеальную программу экспрессии генов в проксимальном отделе тонкой кишки путем ингибирования ацетилирования гистона h4, лизина 27. Biochim Biophys Acta 1839 г., 1273–1282 гг. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Aubin J, Déry U, Lemieux M, Chailler P, Jeannotte L, 2002. Региональная спецификация желудка требует Hoxa5-управляемой мезенхимально-эпителиальной передачи сигналов. Разработка
129, 4075–4087. [PubMed] [Google Scholar]
- Авилион А.А., Николис С.К., Певни Л.Х., Перес Л., Вивиан Н., Ловелл-Бэдж Р., 2003. Мультипотентные клеточные линии в раннем развитии мышей зависят от функции SOX2. Гены Дев 17, 126–140. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ayabe T, Satchell DP, Wilson CL, Parks WC, Selsted ME, Ouellette AJ, 2000. Секреция микробицидных альфа-дефенсинов кишечными клетками Панета в ответ на бактерии. Нат Иммунол 1, 113–118. [PubMed] [Google Scholar]
- Babeu JP, Darsigny M, Lussier CR, Boudreau F, 2009. Ядерный фактор гепатоцитов 4альфа вносит вклад в фенотип кишечного эпителия in vitro и частично играет роль в дифференцировке кишечного эпителия мышей. Am J Physiol Gastrointest Liver Physiol 297, Г124–34. [PubMed] [Google Scholar]
- Bai YQ, Yamamoto H, Akiyama Y, Tanaka H, Takizawa T, Koike M, Kenji Yagi O, Saitoh K, Takeshita K, Iwai T, Yuasa Y, 2002. Эктопическая экспрессия гомеодоменового белка CDX2 при кишечной метаплазии и карциномах желудка. Рак Летт
176, 47–55. [PubMed] [Google Scholar]
- Bass AJ, Watanabe H, Mermel CH, Yu S, Perner S, Verhaak RG, Kim SY, Wardwell L, Tamayo P, Gat-Viks I, Ramos AH, Woo MS, Weir BA, Гетц Г., Бероухим Р., О’Келли М., Датт А., Розенблатт-Розен О., Дзюнич П., Комисаров Дж., Кириак Л.Р., Лафарг С.Дж., Шебл В., Уилбертц Т., Ма К., Рао С., Накагава Х., Лестница Д.Б., Лин Л., Джордано Т.Дж., Вагнер П., Минна Д.Д., Газдар А.Ф., Чжу К.К., Броз М.С., Чекконелло И., Рибейро У., Мари С.К., Даль О., Шивдасани Р.А., Цао М.С., Рубин М.А., Вонг К.К., Регев А., Хан В.К., Пиво Д.Г., Рустги А.К., Мейерсон М., 2009 г.. SOX2 является амплифицированным онкогеном выживания линии при плоскоклеточном раке легкого и пищевода. Нат Жене 41, 1238–1242. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Battle MA, Bondow BJ, Iverson MA, Adams SJ, Jandacek RJ, Tso P, Duncan SA, 2008.
GATA4 необходим для функции тощей кишки у мышей. Гастроэнтерология
135, 1676–1686.e1. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Battle MA, Konopka G, Parviz F, Gaggl AL, Yang C, Sladek FM, Duncan SA, 2006. Ядерный фактор гепатоцитов 4альфа управляет экспрессией белков клеточной адгезии во время эпителиальной трансформации развивающейся печени. Proc Natl Acad Sci U S A 103, 8419–8424. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Beck F, Chawengsaksophak K, Waring P, Playford RJ, Furness JB, 1999. Перепрограммирование кишечной дифференцировки и интеркалярной регенерации у мышей с мутацией Cdx2. Proc Natl Acad Sci U S A 96, 7318–7323. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Бек Ф., Эрлер Т., Рассел А., Джеймс Р., 1995. Экспрессия Cdx-2 в эмбрионе мыши и плаценте: возможная роль в формировании паттерна внеэмбриональных мембран. Дев Дин 204, 219–227. [PubMed] [Google Scholar]
- Beuling E, Aronson BE, Tran LM, Stapleton KA, ter Horst EN, Vissers LA, Verzi MP, Krasinski SD, 2012. GATA6 необходим для пролиферации, миграции, созревания секреторных клеток и экспрессии генов в зрелой толстой кишке мыши. Мол Селл Биол
32, 3392–3402. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Beuling E, Baffour-Awuah NY, Stapleton KA, Aronson BE, Noah TK, Shroyer NF, Duncan SA, Fleet JC, Krasinski SD, 2011. Факторы GATA регулируют пролиферацию, дифференцировку и экспрессию генов в тонком кишечнике половозрелых мышей. Гастроэнтерология 140, 1219–1229.e1. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Beuling E, Bosse T, aan de Kerk DJ, Piaseckyj CM, Fujiwara Y, Katz SG, Orkin SH, Grand RJ, Krasinski SD, 2008. GATA4 опосредует репрессию гена в тонком кишечнике зрелых мышей посредством взаимодействия с кофакторами друга GATA (FOG). Дев Биол 322, 179–189. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Beuling E, Kerkhof IM, Nicksa GA, Giuffrida MJ, Haywood J, aan de Kerk DJ, Piaseckyj CM, Pu WT, Buchmiller TL, Dawson PA, Krasinski SD, 2010 .
Условная делеция Gata4 у мышей индуцирует абсорбцию желчных кислот в проксимальном отделе тонкой кишки. кишки
59, 888–895. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Binder HJ, Reuben A, 2005. Переваривание и всасывание питательных веществ. Медицинская физиология: клеточный и молекулярный подход. 947–974. [Google Scholar]
- Birchenough GM, Johansson ME, Gustafsson JK, Bergström JH, Hansson GC, 2015. Новые разработки в области секреции и функции слизи бокаловидных клеток. Иммунол слизистых оболочек 8, 712–719. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Bjerknes M, Cheng H, 2010. Метастабильность клеточной линии в эпителии кишечника мышей с дефицитом Gfi1. Дев Биол 345, 49–63. [PubMed] [Google Scholar]
- Bosse T, Piaseckyj CM, Burghard E, Fialkovich JJ, Rajagopal S, Pu WT, Krasinski SD, 2006. Gata4 необходим для поддержания идентичности тощей и подвздошной кишке в тонком кишечнике взрослых мышей. Мол Селл Биол 26, 9060–9070. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Castillo D, Puig S, Iglesias M, Seoane A, de Bolós C, Munitiz V, Parrilla P, Comerma L, Poulsom R, Krishnadath KK, Grande L, Pera M , 2012. Активация пути BMP4 и ранняя экспрессия CDX2 характеризуют неспециализированную столбчатую метаплазию в человеческой модели пищевода Барретта. J Gastrointest Surg
16, 227–37; обсуждение 237. [PubMed] [Google Scholar]
- Chawengsaksophak K, de Graaff W, Rossant J, Deschamps J, Beck F, 2004. Cdx2 необходим для осевого удлинения в развитии мышей. Proc Natl Acad Sci U S A 101, 7641–7645. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Chawengsaksophak K, James R, Hammond VE, Köntgen F, Beck F, 1997. Гомеоз и опухоли кишечника у мышей с мутацией Cdx2. Природа 386, 84–87. [PubMed] [Google Scholar]
- Chen C, Fang R, Chou LC, Lowe AW, Sibley E, 2012a. Регуляция PDX1 FABP1 и новых генов-мишеней в эпителиальных клетках кишечника человека Caco-2. Biochem Biophys Res Commun 423, 183–187. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Чен С., Фан Р., Дэвис С., Маравелиас С., Сибли Э., 2009 г.
Инактивация Pdx1, ограниченная эпителием кишечника у мышей, изменяет экспрессию генов двенадцатиперстной кишки в энтероцитах и энтероэндокринных клетках. Am J Physiol Gastrointest Liver Physiol
297, Г1126–37. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Chen C, Leavitt T, Sibley E, 2012b. Кишечный Pdx1 опосредует сети генов метаболизма питательных веществ, и материнская экспрессия необходима для перинатального роста у мышей. Biochem Biophys Res Commun 424, 549–553. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Chen X, Qin R, Liu B, Ma Y, Su Y, Yang CS, Glickman JN, Odze RD, Shaheen NJ, 2008. Многослойный эпителий в модели крысы и пищевода Барретта человека: сходные паттерны экспрессии факторов транскрипции и маркеров дифференцировки. БМК Гастроэнтерол 8, 1. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Cheng H, Leblond CP, 1974. Происхождение, дифференцировка и обновление четырех основных типов эпителиальных клеток в тонкой кишке мыши. V. Унитарная теория происхождения четырех типов эпителиальных клеток. Ам Дж Анат 141, 537–561. [PubMed] [Академия Google]
- Чин А.М., Хилл Д.Р., Аврора М. , Спенс М.Р., 2017.
Морфогенез и созревание эмбрионального и постнатального кишечника. Semin Cell Dev Биол
66, 81–93. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Choi E, Roland JT, Barlow BJ, O’Neal R, Rich AE, Nam KT, Shi C, Goldenring JR, 2014. Атлас распределения клеточных линий желудка человека показывает гетерогенные популяции желез в антральном отделе желудка. кишки 63, 1711–1720 гг. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Чой М.Ю., Ромер А.И., Ван Ю., Ву М.П., Ито С., Лейтер А.Б., Шивдасани Р.А., 2008. Потребность в ограниченном тканями гомеодоменном транскрипционном факторе Nkx6.3 для дифференцировки G-клеток, продуцирующих гастрин, в антральном отделе желудка. Мол Селл Биол 28, 3208–3218. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Даниэли Ю., Ляо Г., Диксон Д., Линнойла Р.И., Лори А., Рэнделл С.Х., Орен М., Джеттен А.М., 2004.
Критическая роль р63 в развитии нормального эпителия пищевода и трахеобронхов. Am J Physiol Cell Physiol
287, C171–81. [PubMed] [Google Scholar]
- Darsigny M, Babeu JP, Dupuis AA, Furth EE, Seidman EG, Lévy E, Verdu EF, Gendron FP, Boudreau F, 2009. Потеря гепатоцитарно-ядерного фактора-4альфа влияет на транспорт ионов в толстой кишке и вызывает хроническое воспаление, напоминающее воспалительное заболевание кишечника у мышей. PLoS один 4, е7609. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Davis RA, Attie AD, 2008. Делеция базолатерального транспортера желчных кислот в подвздошной кишке идентифицирует клеточные сторожевые механизмы, которые регулируют пул желчных кислот. проц. Натл. акад. Sci.USA [бесплатная статья PMC] [PubMed] [Google Scholar]
- De Mey JR, Freund JN, 2013. Понимание эпителиального гомеостаза в кишечнике: старое поле битвы идей, недавние открытия и остающиеся споры. Тканевые барьеры 1, e24965. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Домьян Э.Т., Ферретти Э., Трокмортон К., Мишина Ю., Николис С.К., Sun X, 2011.
Передача сигналов через рецепторы BMP способствует дыхательной идентичности в передней кишке посредством репрессии Sox2. Разработка
138, 971–981. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Drewes T, Senkel S, Holewa B, Ryffel GU, 1996. Изоформы ядерного фактора 4 гепатоцитов человека кодируются отдельными и дифференциально экспрессируемыми генами. Мол Селл Биол 16, 925–931. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Du A, McCracken KW, Walp ER, Terry NA, Klein TJ, Han A, Wells JM, May CL, 2012. Arx необходим для нормального развития энтероэндокринных клеток у мышей и людей. Дев Биол 365, 175–188. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Duggan SP, Behan FM, Kirca M, Zaheer A, McGarrigle SA, Reynolds JV, Vaz GM, Senge MO, Kelleher D, 2016. Характеристика геномной сигнатуры, подобной кишечнику, сохраняющейся во время аденокарциногенеза, связанного с болезнью Барретта, показывает опосредованное NR5A2 стимулирование выживания раковых клеток. научный представитель 6, 32638. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Дункан С.А., Манова К. , Чен В.С., Худлесс П., Вайнштейн Д.К., Бачварова Р.Ф., Дарнелл Дж.Е., 1994.
Экспрессия фактора транскрипции HNF-4 во внеэмбриональной энтодерме, кишечнике и нефрогенной ткани развивающегося эмбриона мыши: HNF-4 является маркером первичной энтодермы в имплантирующейся бластоцисте. Proc Natl Acad Sci U S A
91, 7598–7602. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Dusing MR, Florence EA, Wiginton DA, 2001. Pdx-1 необходим для активации in vivo из энхансера, специфичного для двенадцатиперстной кишки. J Биол Хим 276, 14434–14442. [PubMed] [Академия Google]
- Фанг Р., Олдс Л.С., Сибли Э., 2006 г. Пространственно-временные паттерны экспрессии специфичных для кишечника факторов транскрипции во время постнатального развития кишечника мышей. Паттерны экспрессии генов 6, 426–432. [PubMed] [Google Scholar]
- Feng R, Xiao C, Zavros Y, 2012. Роль Sonic Hedgehog как регулятора функции и дифференцировки желудка. Витам Горм 88, 473–489. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Gao N, White P, Kaestner KH, 2009. Установление кишечной идентичности и эпителиально-мезенхимальной передачи сигналов с помощью Cdx2. Ячейка разработчиков
16, 588–599. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Garrison WD, Battle MA, Yang C, Kaestner KH, Sladek FM, Duncan SA, 2006. Ядерный фактор гепатоцитов 4альфа необходим для эмбрионального развития толстой кишки мыши. Гастроэнтерология 130, 1207–1220. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Gerbe F, Jay P, 2016. Клетки кишечного пучка: эпителиальные стражи, связывающие просветные сигналы с иммунной системой. Иммунол слизистых оболочек 9, 1353–1359. [PubMed] [Google Scholar]
- Gerbe F, Legraverend C, Jay P, 2012. Клетки пучка кишечного эпителия: спецификация и функция. Cell Mol Life Sci 69, 2907–2917. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Gerbe F, Sidot E, Smyth DJ, Ohmoto M, Matsumoto I, Dardalhon V, Cesses P, Garnier L, Pouzolles M, Brulin B, Bruschi M, Harcus Y, Циммерманн В.С., Тейлор Н., Майзелс Р. М., Джей П., 2016.
Клетки кишечного эпителиального пучка инициируют иммунитет слизистой оболочки 2 типа к паразитам-гельминтам. Природа
529, 226–230. [PubMed] [Google Scholar]
- Grainger S, Savory JG, Lohnes D, 2010. Cdx2 регулирует формирование паттерна кишечного эпителия. Дев Биол 339, 155–165. [PubMed] [Google Scholar]
- Gregorieff A, Grosschedl R, Clevers H, 2004. Дефекты задней кишки и трансформация желудочно-кишечного тракта у эмбрионов Tcf4(-/-)/Tcf1(-/-). ЭМБО J 23, 1825–1833 гг. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Guz Y, Montminy MR, Stein R, Leonard J, Gamer LW, Wright CV, Teitelman G, 1995. Экспрессия мышиного STF-1, предполагаемого фактора транскрипции гена инсулина, в бета-клетках поджелудочной железы, эпителии двенадцатиперстной кишки и панкреатических экзокринных и эндокринных предшественниках в онтогенезе. Разработка 121, 11–18. [PubMed] [Академия Google]
- Haumaitre C, Barbacci E, Jenny M, Ott MO, Gradwohl G, Cereghini S, 2005. Отсутствие TCF2/vHNF1 у мышей приводит к агенезии поджелудочной железы. Proc Natl Acad Sci U S A
102, 1490–1495. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Haveri H, Westerholm-Ormio M, Lindfors K, Mäki M, Savilahti E, Andersson LC, Heikinheimo M, 2008. Факторы транскрипции GATA-4 и GATA-6 в нормальной и опухолевой слизистой оболочке желудочно-кишечного тракта человека. БМК Гастроэнтерол 8, 9. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Хаякава Ю., Сакитани К., Кониши М., Асфаха С., Ниикура Р., Томита Х., Ренц Б.В., Тейлор Ю., Маккини М., Миддельхофф М., Цзян З., Танака Т., Дубейковская З.А., Ким В., Чен Х., Урбанска А.М., Нагар К., Вестфален К.Б., Кванте М., Лин К.С., Гершон М.Д., Хара А., Чжао К.М., Чен Д., Уортли Д.Л., Койке К., Ван Т.К., 2017. Фактор роста нервов способствует опухолеобразованию желудка посредством аберрантной холинергической передачи сигналов. Раковая клетка 31, 21–34. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Heller RS, Stoffers DA, Hussain MA, Miller CP, Habener JF, 1998. Неправильная экспрессия панкреатического гомеодоменового белка IDX-1 промотором Hoxa-4 связана с агенезией слепой кишки. Гастроэнтерология
115, 381–387. [PubMed] [Google Scholar]
- Holzer P, 2009. Опиоидные рецепторы желудочно-кишечного тракта. Регул Пепт 155, 11–17. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Honing J, Pavlov KV, Meijer C, Smit JK, Boersma-van Ek W, Karrenbeld A, Burgerhof JG, Kruyt FA, Plukker JT, 2014. Потеря экспрессии CD44 и SOX2 коррелирует с плохим прогнозом у пациентов с аденокарциномой пищевода. Энн Сург Онкол 21 Приложение 4, S657–64. [PubMed] [Академия Google]
- Horst D, Gu X, Bhasin M, Yang Q, Verzi M, Lin D, Joseph M, Zhang X, Chen W, Li YP, Shivdasani RA, Libermann TA, 2010. Потребность эпителий-специфического фактора транскрипции Ets Spdef для функции клеток слизистых желез в антральном отделе желудка. J Биол Хим 285, 35047–35055. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Huh WJ, Esen E, Geahlen JH, Bredemeyer AJ, Lee AH, Shi G, Konieczny SF, Glimcher LH, Mills JC, 2010. XBP1 контролирует созревание зимогенных клеток желудка путем индукции MIST1 и расширения шероховатого эндоплазматического ретикулума. Гастроэнтерология
139, 2038–2049 гг. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Hutton E, Paladini RD, Yu QC, Yen M, Coulombe PA, Fuchs E, 1998. Функциональные различия между кератинами многослойного и простого эпителия. Джей Селл Биол 143, 487–499. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Jacobsen CM, Mannisto S, Porter-Tinge S, Genova E, Parviainen H, Heikinheimo M, Adameyko II, Tevosian SG, Wilson DB, 2005. Взаимодействия GATA-4:FOG регулируют развитие желудочного эпителия у мышей. Дев Дин 234, 355–362. [PubMed] [Академия Google]
- Якобсен К.М., Нарита Н., Белинска М., Сайдер А.Дж., Гордон Дж.И., Уилсон Д.Б., 2002. Генетический мозаичный анализ показывает, что GATA-4 необходим для правильной дифференцировки эпителия желудка мыши. Дев Биол 241, 34–46. [PubMed] [Google Scholar]
- Jeejeebhoy KN, 2002. Синдром короткой кишки: диетологический и медицинский подход. CMAJ
166, 1297–1302. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Jenny M, Uhl C, Roche C, Duluc I, Guillermin V, Guillemot F, Jensen J, Kedinger M, Gradwohl G, 2002. Нейрогенин 3 по-разному необходим для спецификации судеб эндокринных клеток в кишечном и желудочном эпителии. ЭМБО J 21, 6338–6347. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Jepeal LI, Fujitani Y, Boylan MO, Wilson CN, Wright CV, Wolfe MM, 2005. Клеточная экспрессия глюкозозависимого инсулинотропного полипептида регулируется фактором транскрипции PDX-1. Эндокринология 146, 383–391. [PubMed] [Google Scholar]
- Jiang M, Li H, Zhang Y, Yang Y, Lu R, Liu K, Lin S, Lan X, Wang H, Wu H, Zhu J, Zhou Z, Xu J, Lee DK , Zhang L, Lee YC, Yuan J, Abrams JA, Wang TC, Sepulveda AR, Wu Q, Chen H, Sun X, She J, Chen X, Que J, 2017.
Переходные базальные клетки в чешуйчато-столбчатом переходе образуют пищевод Барретта. Природа
550, 529–533. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Kaji I, Kaunitz JD, 2017. Люминальное химосенсирование в гастродуоденальной слизистой оболочке. Карр Опин Гастроэнтерол 33, 439–445. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Khurana S, Mills JC, 2010. Развитие и дифференцировка слизистой оболочки желудка. Prog Mol Biol Transl Sci 96, 93–115. [PubMed] [Google Scholar]
- Ким Б.М., Бюхнер Г., Милетич И., Шарп П.Т., Шивдасани Р.А., 2005. Фактор мезенхимальной транскрипции желудка Barx1 специфицирует идентичность желудочного эпителия посредством ингибирования транзиторной передачи сигналов Wnt. Ячейка разработчиков 8, 611–622. [PubMed] [Академия Google]
- Ким Б.М., Милетич И., Мао Дж., МакМахон А.П., Шарп П.А., Шивдасани Р.А., 2007. Независимые функции и механизмы гомеобоксного гена Barx1 в формировании паттерна желудка и селезенки мыши. Разработка 134, 3603–3613. [PubMed] [Google Scholar]
- Kim SK, Hebrok M, Li E, Oh SP, Schrewe H, Harmon EB, Lee JS, Melton DA, 2000. Паттерн рецептора активина в органогенезе передней кишки. Гены Дев
14, 1866–1871 гг. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Kim TH, Shivdasani RA, 2016. Развитие желудка, стволовые клетки и болезни. Разработка 143, 554–565. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Kohlnhofer BM, Thompson CA, Walker EM, Battle MA, 2016. GATA4 регулирует пролиферацию эпителиальных клеток, чтобы контролировать рост и развитие кишечника у мышей. Селл Мол Гастроэнтерол Гепатол 2, 189–209. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Kokubu H, Ohtsuka T, Kageyama R, 2008. Mash2 необходим для развития нейроэндокринных клеток в железистом желудке. Гены Клетки 13, 41–51. [PubMed] [Google Scholar]
- Koster MI, Kim S, Mills AA, DeMayo FJ, Roop DR, 2004. p63 является молекулярным переключателем для инициации программы эпителиальной стратификации. Гены Дев 18, 126–131. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Kuhnert F, Davis CR, Wang HT, Chu P, Lee M, Yuan J, Nusse R, Kuo CJ, 2004. Существенная потребность в передаче сигналов Wnt для пролиферации тонкой и толстой кишки взрослых, выявленная аденовирусной экспрессией Dickkopf-1. Proc Natl Acad Sci U S A
101, 266–271. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Larsson LI, Madsen OD, Serup P, Jonsson J, Edlund H, 1996. Роль панкреатодуоденального гомеобокса 1 в формировании эндокринного паттерна желудка. Мех Дев 60, 175–184. [PubMed] [Академия Google]
- Ларссон Л.И., Сент-Онж Л., Хугард Д.М., Соса-Пинеда Б., Грусс П., 1998. Pax 4 и 6 регулируют развитие эндокринных клеток желудочно-кишечного тракта. Мех Дев 79, 153–159. [PubMed] [Google Scholar]
- Lee CS, Perreault N, Brestelli JE, Kaestner KH, 2002. Нейрогенин 3 необходим для правильной спецификации энтероэндокринных клеток желудка и поддержания идентичности эпителиальных клеток желудка. Гены Дев 16, 1488–1497. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Li X, Udager AM, Hu C, Qiao XT, Richards N, Gumucio DL, 2009. Динамическое формирование привратника: формирование эпителиальной границы между кишечником и желудком на поздних стадиях внутриутробного развития. Дев Дин
238, 3205–3217. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Litingtung Y, Lei L, Westphal H, Chiang C, 1998. Sonic hedgehog необходим для развития переднего отдела кишечника. Нат Жене 20, 58–61. [PubMed] [Google Scholar]
- Liu K, Jiang M, Lu Y, Chen H, Sun J, Wu S, Ku WY, Nakagawa H, Kita Y, Natsugoe S, Peters JH, Rustgi A, Onaitis MW, Kiernan A , Чен Х, Цюй Дж., 2013. Sox2 взаимодействует с опосредованной воспалением активацией Stat3 при злокачественной трансформации базальных клеток-предшественников передней кишки. Клеточная стволовая клетка 12, 304–315. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Lo HG, Jin RU, Sibbel G, Liu D, Karki A, Joens MS, Madison BB, Zhang B, Blanc V, Fitzpatrick JA, Davidson NO, Konieczny SF, Mills JC, 2017.
Одного фактора транскрипции достаточно, чтобы индуцировать и поддерживать архитектуру секреторных клеток. Гены Дев
31, 154–171. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Madison BB, Braunstein K, Kuizon E, Portman K, Qiao XT, Gumucio DL, 2005. Эпителиальные сигналы hedgehog формируют ось кишечных крипт-ворсинок. Разработка 132, 279–289. [PubMed] [Академия Google]
- Маруяма М., Ичисака Т., Накагава М., Яманака С., 2005 г. Различная роль Sox15 и Sox2 в контроле транскрипции в эмбриональных стволовых клетках мыши. J Биол Хим 280, 24371–24379. [PubMed] [Google Scholar]
- McCauley HA, Guasch G, 2015. Три ура бокаловидной клетке: поддержание гомеостаза в эпителии слизистой оболочки. Тренды Мол Мед 21, 492–503. [PubMed] [Google Scholar]
- McCracken KW, Aihara E, Martin B, Crawford CM, Broda T, Treguier J, Zhang X, Shannon JM, Montrose MH, Wells JM, 2017. Wnt/β-catenin способствует спецификации дна желудка у мышей и людей. Природа 541, 182–187. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- McCracken KW, Catá EM, Crawford CM, Sinagoga KL, Schumacher M, Rockich BE, Tsai YH, Mayhew CN, Spence JR, Zavros Y, Wells JM, 2014. Моделирование развития и заболеваний человека в органоидах желудка, полученных из плюрипотентных стволовых клеток. Природа
516, 400–404. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- McCracken KW, Wells JM, 2017. Механизмы развития эмбрионального желудка. Semin Cell Dev Biol [бесплатная статья PMC] [PubMed] [Google Scholar]
- Miller CT, Moy JR, Lin L, Schipper M, Normolle D, Brenner DE, Iannettoni MD, Orringer MB, Beer DG, 2003. Амплификация генов при аденокарциномах пищевода и болезни Барретта с дисплазией высокой степени. Клин Рак Рез 9, 4819–4825. [PubMed] [Google Scholar]
- Миллс А.А., Чжэн Б., Ван XJ, Фогель Х, Руп Д.Р., Брэдли А., 1999. p63 является гомологом p53, необходимым для морфогенеза конечностей и эпидермиса. Природа 398, 708–713. [PubMed] [Google Scholar]
- Moore BD, Jin RU, Lo H, Jung M, Wang H, Battle MA, Wollheim CB, Urano F, Mills JC, 2016.
Транскрипционная регуляция X-Box-связывающего белка 1 (XBP1) ядерным фактором гепатоцитов 4α (HNF4A) имеет жизненно важное значение для функции бета-клеток. J Биол Хим
291, 6146–6157. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Мунера Дж.О., Сундарам Н., Ранкин С.А., Хилл Д., Уотсон С., Маэ М., Валланс Дж.Е., Шройер Н.Ф., Синагога К.Л., Зарзосо-Лакост А., Хадсон Дж.Р., Хауэлл Дж.К., Чатуведи П., Спенс Дж.Р., Шеннон Дж.М., Зорн А.М., Хелмрат М.А., Уэллс Дж.М., 2017. Дифференцировка плюрипотентных стволовых клеток человека в органоиды толстой кишки посредством временной активации передачи сигналов BMP. Клеточная стволовая клетка 21, 51–64.e6. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Mutoh H, Hakamata Y, Sato K, Eda A, Yanaka I, Honda S, Osawa H, Kaneko Y, Sugano K, 2002. Превращение слизистой оболочки желудка в кишечную метаплазию у трансгенных мышей, экспрессирующих Cdx2. Biochem Biophys Res Commun 294, 470–479. [PubMed] [Google Scholar]
- Ной Т.К., Донахью Б., Шройер Н.Ф., 2011. Развитие и дифференцировка кишечника. Разрешение ячейки опыта 317, 2702–2710. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Parviz F, Matullo C, Garrison WD, Savatski L, Adamson JW, Ning G, Kaestner KH, Rossi JM, Zaret KS, Duncan SA, 2003. Ядерный фактор гепатоцитов 4альфа контролирует развитие печеночного эпителия и морфогенез печени. Нат Жене
34, 292–296. [PubMed] [Академия Google]
- Phillips RW, Frierson HF, Moskaluk CA, 2003. Cdx2 как маркер дифференцировки эпителия кишечника в пищеводе. Ам Дж. Сург Патол 27, 1442–1447. [PubMed] [Google Scholar]
- Qi Z, Chen YG, 2015. Регуляция спецификации судьбы стволовых клеток кишечника. Наука Китай Жизнь Наука 58, 570–578. [PubMed] [Google Scholar]
- Que J, Choi M, Ziel JW, Klingensmith J, Hogan BL, 2006. Морфогенез трахеи и пищевода: текущие игроки и новые роли для головы и Bmps. Дифференциация 74, 422–437. [PubMed] [Академия Google]
- Кью Дж., Окубо Т., Голденринг Дж. Р., Нам К. Т., Куротани Р., Морриси Э. Э., Таранова О., Певный Л. Х., Хоган Б. Л., 2007. Множественные дозозависимые роли Sox2 в формировании паттерна и дифференцировке энтодермы передней передней кишки. Разработка 134, 2521–2531. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Raghoebir L, Bakker ER, Mills JC, Swagemakers S, Kempen MB, Munck AB, Driegen S, Meijer D, Grosveld F, Tibboel D, Smits R, Rottier RJ, 2012. SOX2 перенаправляет судьбу развития кишечного эпителия в сторону преждевременного желудочного фенотипа. Дж Мол Селл Биол
4, 377–385. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Рамальо-Сантос М., Мелтон Д.А., МакМахон А.П., 2000. Сигналы Hedgehog регулируют множество аспектов развития желудочно-кишечного тракта. Разработка 127, 2763–2772. [PubMed] [Google Scholar]
- Ramsey VG, Doherty JM, Chen CC, Stappenbeck TS, Konieczny SF, Mills JC, 2007. Для созревания секретирующих слизь желудочных эпителиальных клеток-предшественников в секретирующие пищеварительные ферменты зимогенные клетки требуется Mist1. Разработка 134, 211–222. [PubMed] [Google Scholar]
- Rodriguez P, Da Silva S, Oxburgh L, Wang F, Hogan BL, Que J, 2010. Передача сигналов BMP в развитии пищевода и преджелудка мыши. Разработка 137, 4171–4176. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Романо Р.А., Смолли К., Магроу С., Серна В.А., Курита Т., Рагхаван С., Синха С., 2012.
Мыши с нокаутом ΔNp63 обнаруживают его незаменимую роль в качестве основного регулятора эпителиального развития и дифференцировки. Разработка
139, 772–782. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- San Roman AK, Shivdasani RA, 2011. Границы, соединения и переходы в желудочно-кишечном тракте. Разрешение ячейки опыта 317, 2711–2718. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Schonhoff SE, Giel-Moloney M, Leiter AB, 2004. Миниобзор: Развитие и дифференцировка эндокринных клеток кишечника. Эндокринология 145, 2639–2644. [PubMed] [Google Scholar]
- Senoo M, Pinto F, Crum CP, McKeon F, 2007. p63 Необходим для пролиферативного потенциала стволовых клеток в многослойном эпителии. Клетка 129, 523–536. [PubMed] [Google Scholar]
- Sheaffer KL, Kaestner KH, 2012. Транскрипционные сети в развитии печени и кишечника. Колд Спринг Харб Перспект Биол 4, а008284. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Sherwood RI, Chen TY, Melton DA, 2009. Транскрипционная динамика образования энтодермальных органов. Дев Дин 238, 29–42. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Sherwood RI, Maehr R, Mazzoni EO, Melton DA, 2011. Передача сигналов Wnt специфицирует и формирует паттерн кишечной энтодермы. Мех Дев
128, 387–400. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Shinohara M, Mao M, Keeley TM, El-Zaatari M, Lee HJ, Eaton KA, Samuelson LC, Merchant JL, Goldenring JR, Todisco A, 2010. Передача сигналов костного морфогенетического белка регулирует развитие и пролиферацию эпителиальных клеток желудка у мышей. Гастроэнтерология 139, 2050–2060.e2. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Silberg DG, Sullivan J, Kang E, Swain GP, Moffett J, Sund NJ, Sackett SD, Kaestner KH, 2002. Эктопическая экспрессия Cdx2 индуцирует желудочную кишечную метаплазию у трансгенных мышей. Гастроэнтерология 122, 689–696. [PubMed] [Google Scholar]
- Simons BD, Clevers H, 2011. Стратегии самообновления гомеостатических стволовых клеток во взрослых тканях. Клетка 145, 851–862. [PubMed] [Google Scholar]
- Sousa JF, Nam KT, Petersen CP, Lee HJ, Yang HK, Kim WH, Goldenring JR, 2016. миР-30-HNF4γ и миР-19Регуляторные сети 4-NR2F2 способствуют усилению маркеров метаплазии в желудке. кишки
65, 914–924. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Spence JR, Lauf R, Shroyer NF, 2011. Развитие кишечной энтодермы позвоночных. Дев Дин 240, 501–520. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Spencer-Dene B, Sala FG, Bellusci S, Gschmeissner S, Stamp G, Dickson C, 2006. Развитие желудка зависит от передачи сигналов, опосредованной фактором роста фибробластов 10/рецептором фактора роста фибробластов 2b. Гастроэнтерология 130, 1233–1244. [PubMed] [Академия Google]
- Стрингер Э.Дж., Дулук И., Саанди Т., Дэвидсон И., Бялецка М., Сато Т., Баркер Н., Клеверс Х., Притчард К.А., Уинтон Д.Дж., Райт Н.А., Фройнд Д.Н., Дешам Дж., Бек Ф., 2012. Cdx2 определяет судьбу постнатальной кишечной энтодермы. Разработка 139, 465–474. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Sulahian R, Chen J, Arany Z, Jadhav U, Peng S, Rustgi AK, Bass AJ, Srivastava A, Hornick JL, Shivdasani RA, 2015. SOX15 регулирует транскрипцию в многослойном эпителии человека и в подмножестве аденокарцином пищевода. Селл Мол Гастроэнтерол Гепатол
1, 598–609.e6. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Танака М., Сайто Х., Кусуми Т., Фукуда С., Симояма Т., Сасаки Ю., Суто К., Мунаката А., Кудо Х., 2001. Пространственное распределение и гистогенез колоректальной метаплазии клеток Панета при идиопатическом воспалительном заболевании кишечника. J Гастроэнтерол Гепатол 16, 1353–1359. [PubMed] [Google Scholar]
- Taraviras S, Mantamadiotis T, Dong-Si T, Mincheva A, Lichter P, Drewes T, Ryffel GU, Monaghan AP, Schütz G, 2000. Первичная структура, хромосомное картирование, экспрессия и транскрипционная активность ядерного фактора мышиных гепатоцитов 4gamma. Биохим Биофиз Акта 1490, 21–32. [PubMed] [Google Scholar]
- Taraviras S, Monaghan AP, Schütz G, Kelsey G, 1994. Характеристика мышиного гена HNF-4 и его экспрессии во время эмбриогенеза мыши. Мех Дев 48, 67–79. [PubMed] [Google Scholar]
- Thompson CA, Battle MA, 2014. Фундаментальная наука о развитии тонкого кишечника, в: Gumucio DL, Samuelson LC, Spence JR (Eds.), Translational Gastroenterology: Organogenesis to Disease. John Wiley & Sons, Ltd, стр. 85–97. [Академия Google]
- Томпсон К.А., Войта К., Пулаканти К., Рао С., Доусон П., Баттл М.А., 2017. GATA4 достаточно для установления идентификации тощей и подвздошной кишки в тонкой кишке. Селл Мол Гастроэнтерол Гепатол 3, 422–446. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Tian X, Jin RU, Bredemeyer AJ, Oates EJ, Błazewska KM, McKenna CE, Mills JC, 2010. RAB26 и RAB3D являются прямыми мишенями транскрипции MIST1, которые регулируют созревание экзокринных гранул. Мол Селл Биол 30, 1269–1284. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Тодиско А, 2017. Регуляция желудочной метаплазии, дисплазии и неоплазии с помощью передачи сигналов костного морфогенетического белка. Селл Мол Гастроэнтерол Гепатол 3, 339–347. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- van de Wetering M, Clevers H, 1993. Sox 15, новый член мышиного семейства Sox транскрипционных факторов HMG box. Нуклеиновые Кислоты Res
21, 1669. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- van den Brink GR, 2007. Сигнализация Hedgehog в развитии и гомеостазе желудочно-кишечного тракта. Физиол Преподобный 87, 1343–1375. [PubMed] [Академия Google]
- ван ден Бринк Г.Р., Блюминг С.А., Хардвик Дж.К., Шепман Б.Л., Офферхаус Г.Дж., Келлер Дж.Дж., Нильсен С., Гаффилд В., ван Девентер С.Дж., Робертс Д.Дж., Пеппеленбош М.П., 2004. Indian Hedgehog является антагонистом передачи сигналов Wnt при дифференцировке эпителиальных клеток толстой кишки. Нат Жене 36, 277–282. [PubMed] [Google Scholar]
- Verzi MP, Khan AH, Ito S, Shivdasani RA, 2008. Фактор транскрипции foxq1 контролирует экспрессию гена муцина и содержание гранул в клетках слизистой оболочки желудка мыши. Гастроэнтерология 135, 591–600. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Verzi MP, Stanfel MN, Moses KA, Kim BM, Zhang Y, Schwartz RJ, Shivdasani RA, Zimmer WE, 2009. Роль гомеодоменового фактора транскрипции Bapx1 в развитии дистального отдела желудка мышей. Гастроэнтерология
136, 1701–1710. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Wada R, Yamaguchi T, Tadokoro K, 2005. Метаплазия клеток Панета толстой кишки является предопухолевым состоянием рака толстой кишки или нет. Джей Карсиног 4, 5. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Уокер EM, Томпсон, Калифорния, Батл, Массачусетс, 2014a. GATA4 и GATA6 регулируют цитодифференцировку кишечного эпителия во время развития. Дев Биол 392, 283–294. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Walker EM, Thompson CA, Kohlnhofer BM, Faber ML, Battle MA, 2014b. Характеристика развивающейся тонкой кишки при отсутствии GATA4 или GATA6. Примечания BMC Res 7, 902. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Walton KD, Freddo AM, Wang S, Gumucio DL, 2016a. Генерация кишечной поверхности: увлекательный рассказ. Разработка 143, 2261–2272. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Уолтон К. Д., Колтеруд А., Червински М.Дж., Белл М.Дж., Пракаш А., Кушваха Дж., Гроссе А.С., Шнелл С., Гумуцио Д.Л., 2012.
Hedgehog-отзывчивые мезенхимальные кластеры направляют формирование паттерна и появление кишечных ворсинок. Proc Natl Acad Sci U S A
109, 15817–15822. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Уолтон К.Д., Уидден М., Колтеруд А., Шоффнер С.К., Червински М.Дж., Кушваха Дж., Пармар Н., Чандхрасекар Д., Фреддо А.М., Шнелл С., Гумуцио Д.Л., 2016b. Виллификация у мышей: сигналы Bmp контролируют формирование паттерна кишечных ворсинок. Разработка 143, 427–436. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ван Д.Х., Тивари А., Ким М.Е., Клемонс Н.Дж., Регми Н.Л., Ходжес В.А., Берман Д.М., Монтгомери Э.А., Уоткинс Д.Н., Чжан С., Чжан К., Цзе С., Спехлер С.Дж., Соуза Р.Ф., 2014. Передача сигналов Hedgehog регулирует FOXA2 в эмбриогенезе пищевода и метаплазии Барретта. Джей Клин Инвест 124, 3767–3780. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Wang X, Ouyang H, Yamamoto Y, Kumar PA, Wei TS, Dagher R, Vincent M, Lu X, Bellizzi AM, Ho KY, Crum CP, Xian W, МакКеон Ф, 2011. Остаточные эмбриональные клетки как предшественники барреттоподобной метаплазии. Клетка
145, 1023–1035. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ван З., Фанг Р., Олдс Л.С., Сибли Э., 2004 г. Транскрипционная регуляция промотора лактазно-флоризингидролазы с помощью PDX-1. Am J Physiol Gastrointest Liver Physiol 287, Г555–61. [PubMed] [Google Scholar]
- Willet SG, Mills JC, 2016. Дифференциация органов желудка и клеток: от эмбриогенеза до гомеостаза взрослых. Селл Мол Гастроэнтерол Гепатол 2, 546–559. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Woo J, Miletich I, Kim BM, Sharpe PT, Shivdasani RA, 2011. Barx1-опосредованное ингибирование передачи сигналов Wnt в передней кишке грудного отдела мыши контролирует трахеопищеводную перегородку и дифференцировку эпителия. PLoS один 6, е22493. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Yang A, Schweitzer R, Sun D, Kaghad M, Walker N, Bronson RT, Tabin C, Sharpe A, Caput D, Crum C, McKeon F, 1999 .